Kio estas metabolo?

Metabolismo aŭ interŝanĝo de substancoj - Aro da kemiaj reagoj okazantaj en vivanta organismo por vivteni la vivon. Ĉi tiuj procezoj permesas al organismoj kreski kaj multigi, konservi siajn strukturojn kaj respondi al mediaj influoj.
Metabolismo estas kutime dividita en 2 stadiojn: katabolismo kaj anabolismo. Dum katabolismo, kompleksaj organikaj substancoj degelas al pli simplaj, kutime liberigante energion. Kaj en la procezoj de anabolismo - de pli simplaj sintezas pli kompleksaj substancoj kaj tio estas akompanata de energiaj kostoj.
Serio de kemiaj metabolaj reagoj nomiĝas metabolaj vojoj. En ili, kun partopreno de enzimoj, iuj biologie signifaj molekuloj estas sinsekve transformitaj al aliaj.
Enzimoj ludas gravan rolon en metabolaj procezoj ĉar:
- agas kiel biologiaj kataliziloj kaj reduktas la aktivigan energion de kemia reago,
- permesas reguligi metabolajn vojojn responde al ŝanĝoj en la medio de la ĉelo aŭ signaloj de aliaj ĉeloj.
Metabolaj ecoj influas ĉu aparta molekulo taŭgas por uzo de la korpo kiel energifonto. Ekzemple, iuj prokariotoj uzas hidrogenan sulfidon kiel energifonton, sed ĉi tiu gaso estas toksa por bestoj. La metabolan indicon ankaŭ influas la kvanto da manĝaĵoj bezonataj por la korpo.
Biologiaj molekuloj
La ĉefaj metabolaj vojoj kaj iliaj komponentoj estas samaj por multaj specioj, kio indikas la unuecon de deveno de ĉiuj vivaj estaĵoj. Ekzemple, iuj karboxilaj acidoj, kiuj estas intermediaj en la ciklo tricarboxilika acido, ĉeestas en ĉiuj organismoj, de bakterioj ĝis eŭkariotaj pluricelaj organismoj. La similecoj en metabolo probable rilatas al la alta efikeco de la metabolaj vojoj, same kiel al ilia frua apero en la historio de evoluo.
Biologiaj molekuloj

Organikaj substancoj, kiuj konsistigas ĉiujn vivajn estaĵojn (bestoj, plantoj, fungoj kaj mikroorganismoj) estas ĉefe reprezentataj de aminoacidoj, karbonhidratoj, lipidoj (ofte nomataj grasoj) kaj nukleaj acidoj. Pro tio ke ĉi tiuj molekuloj estas esencaj por vivo, metabolaj reagoj estas koncentritaj al krei ĉi tiujn molekulojn konstruante ĉelojn kaj histojn aŭ detrui ilin por esti uzataj kiel energifonto. Multaj gravaj biokemiaj reagoj kombiniĝas por sintezi DNA kaj proteinojn.
| Speco de molekulo | Nomo de Formo Monomero | La nomo de la formo de polimero | Ekzemploj de formoj de polimeroj |
|---|---|---|---|
| Aminoacidoj | Aminoacidoj | Proteinoj (polipeptidoj) | Fibrilaj proteinoj kaj globulaj proteinoj |
| Karbonhidratoj | Monosakaridoj | Polisakaridoj | Amido, glukogeno, celulozo |
| Nukleaj acidoj | Nukleotidoj | Polinucleotidoj | ADN kaj RNA |
Metabola rolo
Metabolo meritas esti atentita. Post ĉio, la provizado de niaj ĉeloj kun utilaj substancoj dependas de lia establita laboro. La bazo de metabolo estas kemiaj reagoj, kiuj okazas en la homa korpo. La substancoj necesaj por la vivo de la korpo, kiun ni ricevas per manĝaĵo.
Krome ni bezonas pli da oksigeno, kiun ni spiras kune kun aero. Ideale, ekvilibro estu observata inter la procezoj de konstruado kaj kadukiĝo. Tamen ĉi tiu ekvilibro ofte povas esti ĝenita kaj ekzistas multaj kialoj por tio.
Kaŭzoj de metabolaj malordoj
Inter la unuaj kaŭzoj de metabolaj malordoj povas esti identigita hereda faktoro. Kvankam ĝi estas nekoruptebla, eblas kaj necesas kontraŭbatali ĝin! Ankaŭ metabolaj malordoj povas esti kaŭzitaj de organikaj malsanoj. Tamen ofte ĉi tiuj malordoj estas la rezulto de nia subnutrado.
Kiel superegeco de nutraĵoj, kaj ilia manko tre damaĝas nian korpon. Kaj la konsekvencoj povas esti neinversigeblaj. Troo de certaj nutraĵoj ekestas rezulte de troa konsumo de grasaj manĝaĵoj, kaj manko ekestas el strikta observado de diversaj dietoj por perdo de pezo. La ĉefa dieto plej ofte estas monotona dieto, kio kaŭzas mankon de esencaj nutraĵoj, siavice, tio neeviteble kondukos al disvolviĝo de diversaj malsanoj. Alergio al plej multaj manĝaĵoj eblas.
Metabolaj malsanoj
Eĉ post ekvilibrigado de ĉiuj metabolaj procezoj kaj provizado de la korpo kun la mankantaj vitaminoj, ni riskas ricevi kelkajn gravajn malsanojn kaŭzitajn de la kadukaj produktoj de niaj ĉeloj. Kadukaj produktoj havas ĉion vivan kaj kreskantan, kaj ĉi tio eble estas la plej danĝera malamiko por nia sano. Alivorte, la korpo devas esti purigita de toksinoj en la tempo, aŭ ili simple komencos veneni ĝin. Restante troe, kadukaj produktoj kaŭzas kronikajn malsanojn kaj malrapidigas la laboron de la tuta organismo.
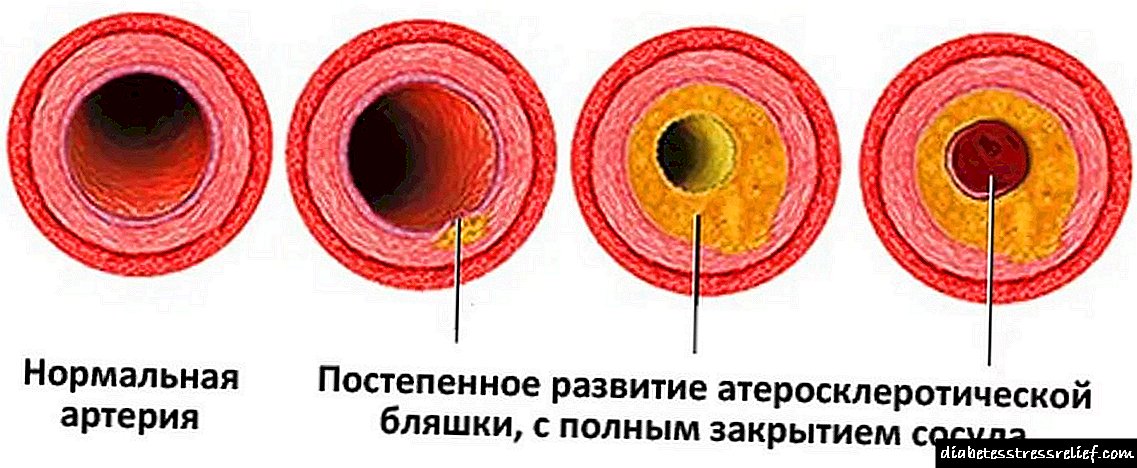
Kun malordoj en la karbonhidrata metabolo, grava malsano okazas - diabeto mellitus, kun netaŭga grasa metabolo, kolesterolo akumuliĝas (Kiel malaltigi kolesterolon hejme sen medikamento?), Kio kaŭzas korajn kaj vaskajn malsanojn. Senpagaj radikaluloj, kiuj pligrandiĝas, kontribuas al apero de malignaj tumoroj.
La obesidad estas ankaŭ komuna konsekvenco de metabolaj problemoj. Ĉi tiu grupo ankaŭ inkluzivas guton, digestajn malordojn, iujn formojn de diabeto ktp. Malekvilibro de mineraloj kaj vitaminoj kondukas al damaĝo de muskoloj, ostoj, severaj malordoj de la kardiovaskula sistemo. En infanoj, ĉi tio povas konduki al tre gravaj konsekvencoj en formo de senĉesa kresko kaj disvolviĝo. Menciindas, ke la plia uzo de vitaminoj ne ĉiam rekomendas, ĉar ilia troa supereco ankaŭ povas havi negativajn konsekvencojn.
Antaŭzorgo
Por reguligi metabolajn procezojn en nia korpo, ni devas scii, ke ekzistas iuj substancoj, kiuj malhelpas la formadon de toksinoj kaj plibonigas la kvaliton de metabolo.
La unua estas oksigeno. La optimuma kvanto da oksigeno en la histoj signife aktivigas metabolajn procezojn.
Due, vitaminoj kaj mineraloj. Kun la aĝo, ĉiuj procezoj malrapidiĝas, parta blokado de sangaj glasoj okazas, do gravas kontroli la ricevon de sufiĉa kvanto da mineraloj, karbonhidratoj kaj oksigeno. Ĉi tio certigos la bonan laboron de la akvo-sala metabolo de la ĉelo, ĉar post la paso de la tempo la ĉelo sekiĝas kaj ne plu ricevas ĉiujn necesajn elementojn por sia vivo. Sciante ĉi, gravas por ni artefarite nutri maljuniĝantajn ĉelojn.
Estas multaj rekomendoj kaj drogoj, kiuj reguligas metabolon. En populara medicino, la algoj de Blanka Maro - fucuso, akiris larĝan popularecon, ĝi enhavas valoran aron da mineraloj kaj utilaj vitaminoj necesaj por plibonigi metabolon. Taŭga nutrado, la ekskludo de la dieto de nutraĵoj enhavantaj kolesterolon kaj aliajn damaĝajn substancojn estas alia maniero por la korpo funkcii senĝene.
Eduko: Moskva Medicina Instituto I. Sechenov, fako - "Medicina komerco" en 1991, en 1993 "Profesiaj malsanoj", en 1996 "Terapio".
Plasta manĝaĵo ujoj: faktoj kaj mitoj!
Aminoacidoj kaj proteinoj Redakti
Proteinoj estas biopolimeroj kaj konsistas el aminoacidaj restaĵoj kunigitaj per peptidaj ligoj. Iuj proteinoj estas enzimoj kaj katalizas kemiajn reagojn. Aliaj proteinoj plenumas strukturan aŭ mekanikan funkcion (ekzemple, formas citoskeleton). Proteinoj ankaŭ ludas gravan rolon en ĉela signalado, imunaj respondoj, ĉela agregado, aktiva transporto trans membranoj kaj regulado de ĉela ciklo.
Kio estas metabolo?

Metabolismo (aŭ metabolo) estas kombinaĵo de la procezoj konverti manĝaĵ-kaloriojn en energion por la vivo de organismo. Metabolismo komenciĝas per digesto kaj fizika aktiveco kaj finiĝas per la spirado de la persono dum dormo, kiam la korpo liveras oksigenon al diversaj organoj sen partopreno de la cerbo kaj tute aŭtonomie.
La koncepto de metabolo tre proksime rilatas al la kalkulo de la ĉiutaga kaloria konsumado, kiu estas la deirpunkto en iu dieto por perdo de pezo aŭ muskola gajno. Surbaze de aĝo, sekso kaj fizikaj parametroj, la nivelo de baza metabolo estas determinita - tio estas la nombro de kalorioj bezonataj por kovri la ĉiutagajn energiajn postulojn de la korpo. Estonte, ĉi tiu indikilo multiplikas per indikilo de homa aktiveco.
Ofte oni kredas, ke rapidigi la metabolon estas bona por perdi pezon, ĉar ĝi kaŭzas, ke la korpo bruligas pli multajn kaloriojn. Reale, la metabolo de perdi pezon homoj kutime malrapidiĝas, ĉar la akcelo de metabolo eblas nur per samtempe pliigo de la kaloria konsumado kaj pliigo de la fizika aktiveco - tio estas dum forto-trejnado por muskola kresko.
Redakti Lipidoj
Lipidoj estas parto de biologiaj membranoj, ekzemple plasmaj membranoj, estas komponantoj de koenzimoj kaj energifontoj. Lipidoj estas hidrofobaj aŭ amfifilaj biologiaj molekuloj, solveblaj en organikaj solviloj kiel benzeno aŭ kloroformo. Grasoj estas granda grupo de komponaĵoj, kiuj inkluzivas grasajn acidojn kaj glicerinon. La glicerol trihidria alkoholo-molekulo, kiu formas tri kompleksajn ester-ligojn kun tri grasaj acidaj molekuloj, estas nomata triglicerido. Kune kun grasaj acidaj restaĵoj, kompleksaj lipidoj povas inkluzivi, ekzemple, sfingosinon (sfingolipidoj), hidrofilajn fosfatajn grupojn (en fosfolipidoj). Esteroidoj, kiel kolesterolo, estas alia granda klaso de lipidoj.
Karbonhidratoj Redakti
Sukeroj povas ekzisti cirkla aŭ lineara formo en la aldehidoj aŭ cetonoj, ili havas plurajn hidroksilajn grupojn. Karbonhidratoj estas la plej oftaj biologiaj molekuloj. Karbonhidratoj plenumas la jenajn funkciojn: konservado kaj transportado de energio (amelo, glukogeno), struktura (planto celulozo, kitino en fungoj kaj bestoj). La plej oftaj sukero-monomeroj estas deksesoj - glukozo, fruktozo kaj galaktozo. Monosakaridoj estas parto de pli kompleksaj liniaj aŭ branĉaj polisakaridoj.
Kiel akceli la metabolon?
La influo de nutrado sur la akcelo de metabolo ne estas tiel klara, kiel ĝi unuavide rigardas. Malgraŭ la fakto, ke ekzistas multaj produktoj, kiuj plimalbonigas metabolon - de tiuj, kiuj kondukas al pezo en sukero kaj aliaj rapidaj karbonhidratoj, al margarino kun ĝiaj trans-grasoj - nur tre malmultaj produktoj povas akceli metabolon.
Ĉar la metabola ciklo de la korpo povas daŭri plurajn tagojn (ekzemple, kun kompleta malakcepto de karbonhidratoj, la korpo ŝanĝos al ketogena dieto nur dum 2-3 tagoj), metabolo ne povas esti akcelita manĝante ununuran produkton aŭ trinkante legoman smoothie por perdi pezon. Inter aliaj aferoj, la akcelo de metabolo kutime asocias kun pliigita apetito - kio ne ĉiam utilas dum sekvo de dieto por perdi pezon.
Metabolaj procezoj de perdo de pezo
Supozu, ke superpezulo decidis perdi pezon, aktive okupiĝis pri fizikaj ekzercoj kaj komencis dieton kun malpliigitaj kalorioj. Li ankaŭ legis, ke por rapidigi la metabolon, vi devas trinki pli da akvo kaj manĝi ananason, riĉan je la enzima bromelino "detruanta grason". Tamen la fina rezulto tute ne estos akcelo de la metabolo, sed ĝia akra malrapidiĝo.
La kialo estas simpla - la korpo komencos sendi signalojn, ke la nivelo de fizika aktiveco kreskis draste, kaj la konsumado de energio de manĝaĵoj malpliiĝis akre. Kaj ju pli aktiva homo okupiĝas pri ekzercoj kaj ju pli strikta dieto observas, des pli forta la korpo pensos, ke venis "malbonaj tempoj" kaj estas tempo malrapidigi la metabolon por ŝpari grasajn rezervojn - krome, kortisolo kaj leptina nivelo kreskos.
Kiel akceli la metabolon?

Por perdi pezon, vi ne bezonas provi "disvastigi" la metabolon kaj rapidigi la metabolon laŭeble. Unue, vi devas pli zorgi pri kiuj produktoj ricevas ĉiutage kaloriojn. Plejofte, normaligo de la dieto kaj kontrolo de glicemia indico de karbonhidratoj konsumitaj rapide kondukos al normaligo de metabolaj procezoj.
Ofte homoj, kiuj provas perdi pezon, superas la energi-kostojn de fizika trejnado, kvankam signife subtaksas la kalorian enhavon de la manĝaĵo, kiun ili konsumas. Ekzemple, la sukero enhavita en unu lato da kolao sufiĉas por daŭro de 30-40 minutoj - alivorte, estas multe pli facile rezigni pri kolao ol elĉerpi vin per elĉerpaj ekzercoj, provi bruligi ĉi tiujn kaloriojn.
Nukleotidoj Redakti
Polimeraj DNA kaj RNA-molekuloj estas longaj, nerangitaj ĉenoj de nukleotidoj. Nukleaj acidoj plenumas la funkcion de stokado kaj efektivigo de genetikaj informoj efektivigitaj dum la procezoj de replikado, transskribo, tradukado kaj proteina biosintezo. La informoj koditaj en nukleaj acidoj estas protektataj kontraŭ ŝanĝoj per sistemoj de riparo kaj multiplikas per repliko de DNA.
Iuj virusoj havas genon enhavantan RNA. Ekzemple, la homa imunodeficacia viruso uzas inversan transskribon por krei DNA-ŝablonon el sia propra genoma RNA. Iuj RNA-molekuloj havas katalizajn proprietojn (ribozimoj) kaj estas parto de spliceosomoj kaj ribosomoj.
Nukleosidoj estas produktoj de aldono de nitrogenaj bazoj al ribosa sukero. Ekzemploj de nitrogenaj bazoj estas heterokiclaj enhavantaj nitrogenoj - derivaĵoj de purinoj kaj pirimidinoj. Iuj nukleotidoj ankaŭ agas kiel koenzimoj en funkciaj grupaj reagoj.
Redakti Koenzimojn
Metabolismo inkluzivas larĝan gamon de kemiaj reagoj, plej multaj rilataj al pluraj ĉefaj specoj de funkciaj grupaj reagaj reagoj. Koenzimoj estas uzataj por translokigi funkciajn grupojn inter enzimoj, kiuj katalizas kemiajn reagojn. Ĉiu klaso de kemiaj reagoj de la translokigo de funkciaj grupoj estas katalizitaj de individuaj enzimoj kaj iliaj kaktaktoroj.
Adenosina trifosfato (ATP) estas unu el la centraj koenzimoj, universala fonto de ĉela energio. Ĉi tiu nukleoto estas uzata por translokigi kemian energion stokitan en makroergiaj ligoj inter diversaj kemiaj reagoj. En ĉeloj, ekzistas malgranda kvanto de ATP, konstante regenerita de ADP kaj AMP. La homa korpo konsumas ATP-mason tage egala al la maso de sia propra korpo. ATP agas kiel ligilo inter katabolismo kaj anabolismo: kun katabolikaj reagoj, ATP formiĝas, kun anabolikaj reagoj, energio konsumas. ATP ankaŭ agas kiel donacanto de la fosfat-grupo en fosforilaj reagoj.
Vitaminoj estas organikaj substancoj kun malalta molekula pezo, kiuj estas necesaj en malgrandaj kvantoj, kaj, ekzemple, ĉe homoj, plej multaj vitaminoj ne estas sintezitaj, sed estas akiritaj per manĝaĵo aŭ per la gastrointestina mikroflora. En la homa korpo, plej multaj vitaminoj estas kaktoj de enzimoj. Plej multaj vitaminoj akiras ŝanĝitan biologian aktivecon, ekzemple, ĉiuj hidrosolvaj vitaminoj en ĉeloj estas fosforiligitaj aŭ kombinitaj kun nukleotidoj. Nicotinamida adenina dinucleotido (NADH) estas derivaĵo de vitamino B3 (niacin), kaj estas grava koenzimo - hidrogena akceptanto. Centoj da malsamaj dehidrogenase-enzimoj forprenas elektronojn de la molekuloj de la substratoj kaj transdonas ilin al la molekuloj NAD +, reduktante ĝin al NADH. La oksidita formo de koenzimo estas substrato por diversaj reduktasoj en la ĉelo. NAD en la ĉelo ekzistas en du rilataj formoj de NADH kaj NADPH. NAD + / NADH estas pli grava por katabolikaj reagoj, kaj NADP + / NADPH estas pli ofte uzata en anabolaj reagoj.
Neorganikaj Substancoj kaj Kofaktoroj Redaktas
Neorganikaj elementoj ludas gravegan rolon en metabolo. Ĉirkaŭ 99% de la maso de mamulo konsistas el karbono, nitrogeno, kalcio, natrio, magnezio, kloro, kalio, hidrogeno, fosforo, oksigeno kaj sulfuro. Biologie signifaj organikaj komponaĵoj (proteinoj, grasoj, karbonhidratoj kaj nukleaj acidoj) enhavas grandan kvanton da karbono, hidrogeno, oksigeno, nitrogeno kaj fosforo.
Multaj neorganikaj komponaĵoj estas ionikaj elektrolitoj. La plej gravaj jonoj por la korpo estas natrio, kalio, kalcio, magnezio, kloridoj, fosfatoj kaj bicarbonatoj. La ekvilibro de ĉi tiuj jonoj ene de la ĉelo kaj en la eksterĉela mezuro determinas la osmotan premon kaj pH. Ionaj koncentriĝoj ankaŭ ludas gravan rolon en la funkciado de nervaj kaj muskolaj ĉeloj. La agada potencialo en ekscitaj histoj ekestas el interŝanĝo de jonoj inter la eksterĉela fluido kaj la citoplasmo. Elektrolitoj eniras kaj eliras el la ĉelo tra jonoj-kanaloj en la plasmembrano. Ekzemple, dum muskola kuntiriĝo, kalcioj, natrio, kaj kalio jonoj moviĝas en la plasmembranon, citoplasmon kaj T-tubojn.
Transiraj metaloj en la korpo estas spuroj, zinko kaj fero estas la plej oftaj. Ĉi tiuj metaloj estas uzataj de iuj proteinoj (ekzemple enzimoj kiel koaktoroj) kaj gravas por reguligi la agadon de enzimoj kaj transportaj proteinoj. Kofaktoroj de enzimoj kutime forte ligiĝas al specifa proteino, tamen ili povas esti modifitaj dum katalizo, kaj post katalizo ili ĉiam revenas al sia originala stato (ne konsumatas). Spuroj de metaloj estas sorbitaj de la korpo uzante specialajn transportajn proteinojn kaj ne troviĝas en la korpo en libera stato, ĉar ili estas asociitaj kun specifaj portantaj proteinoj (ekzemple feritino aŭ metalotioneinoj).
Ĉiuj vivantaj organismoj povas esti dividitaj en ok ĉefajn grupojn, laŭ kiuj oni uzas ĝin: energifonto, karbona fonto kaj donanto de elektronoj (oksidigebla substrato).
- Kiel fonto de energio, vivantaj organismoj povas uzi: la energio de lumo (foto) aŭ la energio de kemiaj ligoj (moemo) Aldone, por priskribi parazitajn organismojn per la energiaj rimedoj de la gastiganta ĉelo, la termino paratrofo.
- Kiel donacanto de elektronoj (reduktanta agento), vivantaj organismoj povas uzi: neorganikaj substancoj (rolantaro) aŭ organika materio (organo).
- Kiel karbona fonto, vivaj organismoj uzas: karbondioksido (aŭtomobilo) aŭ organika materio (hetero-) Iafoje terminoj aŭtomobilo kaj heterotrofo uzata rilate al aliaj elementoj, kiuj formas parton de biologiaj molekuloj en reduktita formo (t.e. nitrogeno, sulfuro). En ĉi tiu kazo, "nitrogen-aŭtotrofaj" organismoj estas specioj, kiuj uzas oksidajn neorganikajn komponaĵojn kiel nitrogenan fonton (ekzemple plantoj povas efektivigi reduktadon de nitratoj). Kaj "nitrogenaj heterotrofoj" estas organismoj nekapablaj efektivigi la redukton de oksidigitaj formoj de nitrogeno kaj uzi organikajn komponaĵojn kiel ties fonton (ekzemple, bestoj por kiuj aminoacidoj estas la fonto de nitrogeno).
La nomo de la tipo de metabolo formiĝas per aldono de la respondaj radikoj kaj aldono ĉe la fino de la radiko -trofo-. La tablo montras la eblajn specojn de metabolo per ekzemploj:
| Fonto energio | Donacanto de elektronoj | Karbonfonto | Tipo de metabolo | Ekzemploj |
|---|---|---|---|---|
| Sunbrilo Foto | Organika materio organo | Organika materio heterotrofo | Foto-heterotrofaj organoj | Purpuraj ne-sulvaj bakterioj, halobakterioj, Iuj cianobakterioj. |
| Karbona dioksido aŭtotrofo | Foto-organotrofoj | Malofta speco de metabolo asociita kun la oksidado de nedifekteblaj substancoj. Ĝi estas karakteriza por iuj purpuraj bakterioj. | ||
| Neorganikaj substancoj rolantaro* | Organika materio heterotrofo | Foto de litiaj heterotrofoj | Iuj cianobakterioj, purpuraj kaj verdaj bakterioj, ankaŭ estas heliobakterioj. | |
| Karbona dioksido aŭtotrofo | Foto litototofoj | Pli altaj plantoj, algoj, cianobakterioj, purpuraj sulfaj bakterioj, verdaj bakterioj. | ||
| La energio kemia ligoj Kemo- | Organika materio organo | Organika materio heterotrofo | Heterotrofoj de Chemo Organo | Bestoj, Fungoj, Plej multaj mikroorganismoj de reduktantoj. |
| Karbona dioksido aŭtotrofo | Hemo-Organotrofoj | Oksidado de malfacilaj asimilaj substancoj, ekzemple laŭvolaj metilotrofoj, oksidanta formika acido. | ||
| Neorganikaj substancoj rolantaro* | Organika materio heterotrofo | Kemio-litiaj heterotrofoj | Metano-formanta arkaon, Hidrogenajn bakteriojn. | |
| Karbona dioksido aŭtotrofo | Kemo-Litotrofoj | Feraj bakterioj, Hidrogenaj bakterioj, Nitrifiaj bakterioj, Serobakterioj. |
- Iuj aŭtoroj uzas -hidro kiam akvo agas kiel elektrono-donacanto.
La klasifiko estis ellaborita de grupo de aŭtoroj (A. Lvov, C. van Nil, F. J. Ryan, E. Tatem) kaj aprobita en la 11a simpozio en laboratorio de Cold Spring Harbor kaj estis uzata origine por priskribi la tipojn de nutrado de mikroorganismoj. Tamen ĝi estas uzata nuntempe por priskribi la metabolon de aliaj organismoj.
Estas evidente de la tabelo, ke la metabolaj kapabloj de procariotoj estas multe pli diversaj kompare al eŭkariotoj, kiuj estas karakterizitaj per la fotolitokaj kaj kemioorganoheterotrofaj tipoj de metabolo.
Oni devas rimarki, ke iuj specoj de mikroorganismoj povas, dependante de mediaj kondiĉoj (lumigado, havebleco de organikaj substancoj, ktp) kaj fiziologia stato, efektivigi metabolon de diversaj specoj. Ĉi tiu kombinaĵo de pluraj specoj de metabolo estas priskribita kiel miksotrofio.
Kiam vi aplikas ĉi tiun klasifikon al pluricelaj organismoj, gravas kompreni, ke en unu organismo povas ekzisti ĉeloj, kiuj diferencas laŭ la tipo de metabolo. Do la ĉeloj de la aeraj, fotosintezaj organoj de plurĉelaj plantoj estas karakterizitaj de la fotolitoŭtotrofa speco de metabolo, dum la ĉeloj de la subteraj organoj estas priskribitaj kiel kemioorganoterotrofaj. Kiel en la kazo de mikroorganismoj, kiam la mediaj kondiĉoj, stadio de evoluo kaj fiziologia stato ŝanĝiĝas, la tipo de metabolo de la ĉeloj de pluricela organismo povas ŝanĝiĝi. Ekzemple, en la mallumo kaj en la stadio de semado de ĝermado, ĉeloj de pli altaj plantoj metaboligas kemio-organo-heterotrofan tipon.
Metabolismo estas nomata metabolaj procezoj, en kiuj disiĝas relative grandaj organikaj molekuloj de sukeroj, grasoj kaj aminoacidoj. Dum katabolismo, pli simplaj organikaj molekuloj formiĝas, kiuj estas necesaj por anabolismo (biosintezo) reagoj. Ofte estas en la kurso de katabolismaj reagoj, ke la korpo mobilizas energion, tradukante la energion de la kemiaj ligoj de organikaj molekuloj akiritaj dum la digesto de manĝaĵoj, en alireblajn formojn: en la formo de ATP, reduktitaj koenzimoj kaj transmembrana elektrokemia. La esprimo katabolismo ne estas sinonimo de "energia metabolo": en multaj organismoj (ekzemple fototrofoj), la ĉefaj procezoj de konservado de energio ne rekte rilatas al la disrompo de organikaj molekuloj. La klasifiko de organismoj laŭ tipo de metabolo povas esti bazita sur la fonto de energio, kiel reflektita en la antaŭa sekcio. Kemotrofoj uzas la energion de kemiaj ligoj, kaj fototrofoj konsumas la energion de sunlumo. Tamen ĉiuj ĉi tiuj malsamaj formoj de metabolo dependas de redoxaj reagoj, kiuj estas asociitaj kun la translokigo de elektronoj de reduktitaj donacantoj de molekuloj, kiel organikaj molekuloj, akvo, amoniako, hidrogena sulfido, al akceptantaj molekuloj kiel oksigeno, nitratoj aŭ sulfato. Ĉe bestoj, ĉi tiuj reagoj implikas rompon de kompleksaj organikaj molekuloj en pli simplajn, ekzemple karbondioksido kaj akvo. En fotosintezaj organismoj - plantoj kaj cianobakterioj - reagoj de elektronaj translokigoj ne liberigas energion, sed ili estas uzataj kiel maniero stoki energion absorbitan de sunlumo.
Katabolismo en bestoj povas esti dividita en tri ĉefajn stadiojn. Unue, grandaj organikaj molekuloj kiel proteinoj, polisakaridoj kaj lipidoj malpliiĝas al pli malgrandaj komponentoj ekster la ĉeloj. Plue, ĉi tiuj malgrandaj molekuloj eniras la ĉelojn kaj transformiĝas en eĉ pli malgrandajn molekulojn, ekzemple, acetil-CoA. Siavice, la acetila grupo de koenzima A oxidas al akvo kaj karbona dioksido en la ciklo de Krebs kaj la spira ĉeno, liberigante energion, kiu estas stokita en la formo de ATP.
Redakta Digesto
Makromolekuloj kiel amelo, celulozo aŭ proteinoj devas diseriĝi al pli malgrandaj ekzempleroj antaŭ ol ili povas esti uzataj de ĉeloj. Pluraj klasoj de enzimoj estas implikitaj en degenero: proteazoj, kiuj detruas proteinojn al peptidoj kaj aminoacidoj, glicozidasojn, kiuj detruas polisakaridojn al oligo- kaj monosakaridoj.
Mikroorganismoj apartigas hidrolitajn enzimojn en la spacon ĉirkaŭ ili, kiu diferencas de bestoj, kiuj sekrecias tiajn enzimojn nur de specialigitaj glandaj ĉeloj. Aminoacidoj kaj monosakaridoj, rezultantaj el la agado de eksterĉelaj enzimoj, tiam eniras la ĉelojn per aktiva transporto.
Akiri Energio Redakti
Dum karbonhidrata katabolismo, kompleksaj sukeroj rompiĝas al monosakaridoj, kiuj estas absorbitaj de ĉeloj. Unufoje enen, sukeroj (ekzemple, glukozo kaj fruktozo) estas konvertitaj al piruvato dum glicolizo, kaj oni produktas certan kvanton da ATP. Piravida acido (piruvato) estas intermediaro en pluraj metabolaj vojoj. La ĉefa vojo de piruvata metabolo estas konvertiĝo al acetil-CoA kaj poste al trikarboxilika acida ciklo. Samtempe, parto de la energio estas stokita en la ciklo de Krebs en la formo de ATP, kaj ankaŭ reestigas NADH kaj FAD-molekulojn. En la procezo de glicolizo kaj la tricarboxilika acida ciklo, formiĝas karbona dioksido, kio estas kromprodukto de la vivo. Sub anaerobiaj kondiĉoj, rezulte de glicolizo el piruvato kun partopreno de la enzimo laktata dehidrogenazo, laktato formiĝas, kaj NADH estas oksidita al NAD +, kiu estas reuzita en glicolizaj reagoj. Ekzistas ankaŭ alternativa vojo por la metabolo de monosakaridoj - la vojo de pentosa fosfato, dum kiu la energio estas stokita en la formo de reduktita koenzima NADPH kaj pentozoj formiĝas, ekzemple, ribose, kio estas necesa por la sintezo de nukleaj acidoj.
Grasoj en la unua etapo de katabolismo estas hidroligitaj en liberajn grasajn acidojn kaj glicerinon. Gravaj acidoj diseriĝas dum beta-oksidado por formi acetil-CoA, kiu plue kataboliĝas en la ciklo de Krebs, aŭ iras al la sintezo de novaj grasaj acidoj. La grasaj acidoj liberigas pli da energio ol karbonhidratoj, ĉar grasoj enhavas specife pli da hidrogenajn atomojn en sia strukturo.
Aminoacidoj estas aŭ uzataj por sintezi proteinojn kaj aliajn biomolekulojn, aŭ estas oksiditaj al ureo, karbona dioksido kaj servas kiel fonto de energio. La oksida vojo de katabolismo de aminoacidoj komenciĝas per la forigo de la amino-grupo per transaminazaj enzimoj. Amino-grupoj estas uzataj en la urea ciklo, aminoacidoj malhavantaj amino-grupojn nomiĝas ketoacidoj. Iuj keto-acidoj estas intermedioj en la ciklo de Krebs. Ekzemple, malaminado de glutamato produktas alfa-ketoglutarikan acidon. Glicogenaj aminoacidoj ankaŭ povas esti konvertitaj al glukozo en reagoj de gluconeogenezo.
Oxidativa fosforilado Redakti
En oxidativa fosforilado, elektronoj forigitaj de manĝaĵaj molekuloj laŭ la metabolaj vojoj (ekzemple en la ciklo de Krebs) estas translokigitaj al oksigeno, kaj la liberigita energio estas uzata por sintezi ATP. En eŭkariotoj, ĉi tiu procezo efektiviĝas kun la partopreno de kelkaj proteinoj fiksitaj en mitokondriaj membranoj, nomataj spira ĉeno de elektron-translokigo. En procariotoj, ĉi tiuj proteinoj ĉeestas en la interna membrano de la ĉela muro. Proteinoj de la elektron-translokiga ĉeno uzas la energion akiritan per translokado de elektronoj de reduktitaj molekuloj (t.e. NADH) al oksigeno por pumpi protonojn tra la membrano.
Kiam oni protektas protonojn, oni kreas diferencon en la koncentriĝo de hidrogenaj jonoj kaj okazas elektrokemia kemia gradiento. Ĉi tiu forto resendas protonojn reen al mitokondrioj tra la bazo de ATP-sintasio. La fluo de protonoj igas la ringon el la c-subunuoj de la enzimo rotacii, rezulte de kio la aktiva centro de sintakso ŝanĝas sian formon kaj fosforiligas adenosinan difosfaton, igante ĝin ATP.
Neorganika Energio Redakti
Hemolitotrofoj estas nomataj prokariotoj, kiuj havas specialan specon de metabolo, en kiu energio formiĝas rezulte de la oksido de neorganikaj komponaĵoj. Kemiolitotrofoj povas oksidi molekulan hidrogenon, sulfurajn komponaĵojn (t.e. sulfidoj, hidrogenan sulfidon kaj neorganikajn tiosulfatojn), feron (II) oksidon aŭ amonion. En ĉi tiu kazo, la energio el la oksido de ĉi tiuj komponaĵoj estas generita de elektronaj akceptiloj, kiel oksigeno aŭ nitrito. La procezoj por akiri energion el neorganikaj substancoj ludas gravan rolon en tiaj biogeokemiaj cikloj kiel acetogenezo, nitrifigado kaj denitrifigado.
Redaktas Energio de Sunlumo
La energio de sunlumo estas sorbita de plantoj, cianobakterioj, purpuraj bakterioj, verdaj sulfaj bakterioj, kaj iuj protozooj. Ĉi tiu procezo ofte kombiniĝas kun la konvertiĝo de karbona dioksido al organikaj komponaĵoj kiel parto de la fotosinteza procezo (vidu sube). La sistemoj de kaptado de energio kaj fija karbono en iuj prokariotoj povas funkcii aparte (ekzemple en purpuraj kaj verdaj sulfaj bakterioj).
En multaj organismoj, la absorbo de sunenergio estas principe simila al oksorga fosforiligo, ĉar en ĉi tiu kazo la energio estas stokita en formo de protona koncentriĝa gradiento kaj la motoro de la protonoj kondukas al la sintezo de ATP. La elektronoj bezonataj por ĉi tiu translokiga ĉeno venas de malpezaj rikoltaj proteinoj nomataj fotosintezaj reagaj centroj (ekzemple rodopsinoj). Depende de la tipo de fotosintezaj pigmentoj, oni klasifikas du specojn de reakciaj centroj; nuntempe, plej multaj fotosintezaj bakterioj havas nur unu tipon, dum plantoj kaj cianobakterioj estas du.
En plantoj, algoj kaj cianobakterioj, fotosistemo II uzas la energion de lumo por forigi elektronojn el akvo, kun molekula oksigeno liberigita kiel kromprodukto de la reago. La elektronoj tiam eniras la kompleksan citokroman b6f, kiu uzas energion por pumpi protonojn tra la tilacoida membrano en kloroplastoj. Sub la influo de elektrokemia gradiento, protonoj moviĝas reen tra la membrano kaj ekigas ATP-sintazon. La elektronoj tiam trapasas fotosistemon I kaj povas esti uzataj por restarigi la NADP + koenzimon, por uzi en la kalvinciklo, aŭ por reciklado por formi pliajn ATP-molekulojn.
Anabolismo - aro de metabolaj procezoj de biosintezo de kompleksaj molekuloj kun elspezo de energio. La kompleksaj molekuloj, kiuj konsistigas la ĉelajn strukturojn, estas sintezitaj sinsekve el pli simplaj pioniroj. Anabolismo inkluzivas tri ĉefajn stadiojn, ĉiu el kiuj estas katalizita de specialigita enzimo. En la unua etapo, pioniraj molekuloj estas sintezitaj, ekzemple, aminoacidoj, monosakaridoj, terpenoidoj kaj nukleotidoj. En la dua etapo, pioniroj per elspezo de ATP-energio konvertiĝas al aktivigitaj formoj. En la tria etapo, la aktivigitaj monomeroj estas kombinitaj en pli kompleksajn molekulojn, ekzemple proteinoj, polisakaridoj, lipidoj kaj nukleaj acidoj.
Ne ĉiuj vivaj organismoj povas sintezi ĉiujn biologie aktivajn molekulojn. Autotrofoj (ekzemple plantoj) povas sintezi kompleksajn organikajn molekulojn el simplaj neorganikaj malalt-molekulaj substancoj kiel karbona dioksido kaj akvo. Heterotrofoj bezonas fonton de pli kompleksaj substancoj, kiel monosakaridoj kaj aminoacidoj, por krei pli kompleksajn molekulojn. Organismoj estas klasifikitaj laŭ siaj ĉefaj energifontoj: fotoautotrofoj kaj fotoheterotrofoj ricevas energion el sunlumo, dum kemioautotrofoj kaj kemiheterotrofoj ricevas energion de neorganikaj oksidaj reagoj.
Eldono pri Karbono-Bindita
Fotosintezo estas la procezo de biosintezo de sukeroj el karbona dioksido, en kiu la necesa energio estas absorbita de sunlumo. En plantoj, cianobakterioj kaj algoj, fotolizo de akvo okazas dum fotosintezo de oksigeno, dum oksigeno estas liberigita kiel kromprodukto. Por konverti CO2 3-fosfoglicerato uzas la energion de ATP kaj NADP stokita en fotosistemoj. La karbona liganta reago efektiviĝas uzante la enzimo ribulosa bisfosfata karboxilazo kaj estas parto de la kalva ciklo. Tri specoj de fotosintezo estas klasifikitaj en plantoj - laŭ la vojo de tri-karbonaj molekuloj, laŭ la vojo de kvar-karbonaj molekuloj (C4), kaj CAM-fotosintezo. Tri specoj de fotosintezo malsamas en la vojo de ligado al karbona dioksido kaj ĝia eniro en la ciklon de Calvin; en C3-plantoj, ligado de CO2 okazas rekte en la kalvina ciklo, kaj ĉe C4 kaj CAM CO2 antaŭe inkluzivita en aliaj komponaĵoj. Malsamaj formoj de fotosintezo estas adaptiĝoj al la intensa fluo de sunlumo kaj al sekaj kondiĉoj.
En fotosintezaj prokariotoj, la mekanismoj de ligado de karbono estas pli diversaj. Karbona dioksido povas esti fiksita en la kalvina ciklo, en la inversa ciklo de Krebs, aŭ en acetil-CoA-karboxilataj reagoj. Prokariotoj - kemioautotrofoj ankaŭ ligas CO2 tra la ciklo de Calvin, sed energio el neorganikaj komponaĵoj estas uzata por efektivigi la reagon.
Karbonhidratoj kaj Glycans Redakti
En la procezo de sukura anabolismo, simplaj organikaj acidoj povas esti konvertitaj al monosakaridoj, ekzemple glukozo, kaj tiam uzataj por sintezi polisakaridojn, kiel amelon. La formado de glukozo el komponaĵoj kiel piruvato, laktato, glicerino, 3-fosfogliccerato kaj aminoacidoj estas nomata gluconeogenezo. En la procezo de gluconeogenezo, piruvato estas konvertita al glukozo-6-fosfato per serio de interaj komponaĵoj, multaj el kiuj ankaŭ formiĝas dum glicolizo. Tamen, glukogenozo ne estas nur glicolizo en la kontraŭa direkto, ĉar pluraj kemiaj reagoj katalizas specialajn enzimojn, kio ebligas sendepende reguligi la procezojn de formado kaj rompo de glukozo.
Multaj organismoj stokas nutraĵojn en formo de lipidoj kaj grasoj, tamen vertebruloj ne havas enzimojn, kiuj katalizas la konvertiĝon de acetil-CoA (produkto de metabolo de grasaj acidoj) al piruvato (substrato de glukogenozo). Post longedaŭra malsato, vertebruloj komencas sintezi korpojn de cetonoj el grasaj acidoj, kiuj povas anstataŭigi glukozon en histoj kiel la cerbo. En plantoj kaj bakterioj, ĉi tiu metabola problemo solviĝas per la ciklo de gloksilato, kiu preterpasas la stadion de malkarboxilado en la ciklo de cítricos kaj permesas konverti acetil-CoA al oksaloacetato, kaj poste uzi ĝin por sintezo de glukozo.
Polisakaridoj plenumas strukturajn kaj metabolajn funkciojn, kaj ankaŭ povas esti kombinitaj kun lipidoj (glicolipidoj) kaj proteinoj (glicoproteinoj) uzante oligosakaridajn transferajn enzimojn.
Gravaj Acidoj, Izoprenoidoj kaj Steroj
La grasaj acidoj estas formitaj de grasaj acidaj sintezoj el acetil-CoA. La karbona skeleto de grasaj acidoj etendiĝas en la ciklo de reagoj, en kiuj unue kuniĝas la acetila grupo, poste la karbonila grupo reduktiĝas al la hidroksil-grupo, poste okazas deshidratado kaj posta resaniĝo. Enzimaj grasaj biosintezaj enzimoj estas klasifikitaj en du grupojn: ĉe bestoj kaj fungoj, ĉiuj reagaj sintezaj grasaj acidoj estas farataj de unu multfunkcia proteino tipo I, en plantaj plastidoj kaj en bakterioj, ĉiu tipo katalizas per apartaj enzimoj de tipo II.
Terpenoj kaj terpenoidoj estas reprezentantoj de la plej granda klaso de herbaj naturaj produktoj. Reprezentantoj de ĉi tiu grupo de substancoj estas derivaĵoj de izopreno kaj formiĝas el aktivigitaj pioniroj de izopentila pirofosfato kaj dimetilalil-pirofosfato, kiuj siavice formiĝas en malsamaj metabolaj reagoj. En bestoj kaj arkaoj, izopentila pirofosfato kaj dimetilalil-pirofosfato estas sintezitaj el acetil-CoA en la mevalonata vojo, dum en plantoj kaj bakterioj, piruvato kaj gliceraldehido-3-fosfato estas substratoj de la ne-mevalonata vojo. En steroidaj biosintezaj reagoj, izoprenaj molekuloj kombinas kaj formas skalenon, kiuj tiam formas ciklajn strukturojn kun la formado de lanosterolo. Lanosterolo povas esti konvertita al aliaj steroidoj, kiel kolesterolo kaj ergosterolo.
Ŝtonetoj Redakti
Organismoj malsamas en sia kapablo sintezi 20 oftajn aminoacidojn. Plej multaj bakterioj kaj plantoj povas sintezi ĉiujn 20, sed mamuloj kapablas sintezi nur 10 esencajn aminoacidojn. Tiel, en la kazo de mamuloj, 9 esencaj aminoacidoj devas esti akiritaj el manĝaĵo. Ĉiuj aminoacidoj estas sintezitaj el intermediaĵoj de glicolizo, ciklo de cítrico acido, aŭ vojo de pentosa monofosfato. La translokigo de amino-grupoj de aminoacidoj al alfa-keto-acidoj nomiĝas transaminado. Donacantoj de amino-grupoj estas glutamato kaj glutamino.
Aminoacidoj ligitaj per peptidaj ligoj formas proteinojn. Ĉiu proteino havas unikan sinsekvon de aminoacidaj restaĵoj (primara proteina strukturo). Same kiel la literoj de la alfabeto povas esti kombinitaj kun formado de preskaŭ senfinaj variaĵoj de vortoj, aminoacidoj povas ligiĝi en unu sinsekvo aŭ alia kaj formi diversajn proteinojn. La aminoacil-tRNA-sintetasa enzimo katalizas ATP-dependan aldonon de aminoacidoj al tRNA kun ester-ligoj, kaj aminoacil-tRNAoj formiĝas. Aminoacil-tRNAoj estas substratoj por ribosomoj, kiuj kombinas aminoacidojn en longajn polipeptidajn ĉenojn per mRNA-matrico.




-
La uzo de la drogo Ranitidine por pancreatito
Ranitidine por recenzoj pri pancreatitis pri la apliko Ni proponas legi la artikolon pri la temo: "ranitidino por recenzoj pri pancreatitis por la apliko" kun komentoj de profesiuloj. ... -
-
-