La rolo de proteino en la homa korpo

La granda graveco de proteinoj por la korpo ŝuldiĝas al iliaj funkcioj.
La prezentitaj bazaj funkcioj de proteinoj ilustras la gravecon de ĉi tiu klaso de substancoj por certigi normalan homan vivon.
En la 19-a jarcento, sciencistoj deklaris:
- proteinaj korpoj estas unikaj, la esenco de vivo,
- konstanta metabolo inter vivuloj kaj la medio estas bezonata.
Ĉi tiuj dispozicioj restas ĝis nun.
La baza konsisto de proteinoj
La grandegaj molekulaj unuoj de simpla proteino nomata proteino estas formitaj de malgrandaj kemie ligitaj blokoj - aminoacidoj kun identaj kaj malsamaj fragmentoj. Tiaj strukturaj komponaĵoj nomiĝas heteropolimeroj. Nur 20 reprezentantoj de la klaso de aminoacidoj ĉiam troviĝas en naturaj proteinoj. La baza konsisto de proteinoj estas karakterizita per la deviga ĉeesto de karbono - C, nitrogeno - N, hidrogeno - H, oksigeno - O. Sulfuro - S. Ofte troveblas En kompleksaj proteinoj nomataj proteidoj, aliaj substancoj enhavas krom aminoacidaj restaĵoj. Laŭe, fosforo - P, kupro - Cu, fero - Fe, jodo - I, kaj seleno - Se povas esti en sia kunmetaĵo.
Aminokarboxilikaj acidoj de naturaj proteinoj estas klasifikitaj laŭ kemia strukturo kaj biologia graveco. Kemia klasifiko gravas por kemiistoj, biologiaj - por ĉiuj.
En la homa korpo ĉiam estas du fluoj de transformoj:
- la rompo, oksido, forigo de manĝaĵoj,
- biologia sintezo de novaj esencaj substancoj.
12 aminoacidoj el ĉiam trovitaj en naturaj proteinoj povas esti kreitaj per biologia sintezo de la homa korpo. Ili estas nomataj interŝanĝeblaj.
8 aminoacidoj neniam estas sintezitaj ĉe homoj. Ili estas nemalhaveblaj, devas esti donitaj regule kun manĝaĵo.
Laŭ ĉeesto de esencaj amino-karboxilaj acidoj, proteinoj dividiĝas en du klasojn.
- Kompletaj proteinoj havas ĉiujn aminoacidojn bezonatajn de la homa korpo. La bezonata aro da esencaj aminoacidoj enhavas proteinojn de dometo, laktaĵoj, birdoj, viandoj de brutoj, maro kaj dolĉakvaj fiŝoj, ovoj.
- En difektaj proteinoj, eble mankas unu aŭ pli gravaj acidoj. Ĉi tiuj inkluzivas plantajn proteinojn.
Por taksi la kvaliton de dietaj proteinoj, la tutmonda medicina komunumo komparas ilin kun la "ideala" proteino, kiu strikte kontrolis proporciojn de esencaj kaj esencaj aminoacidoj. En naturo, "ideala" proteino ne ekzistas. Tiel proksime al li kiel bestaj proteinoj. Plantaj proteinoj ofte ne sufiĉas al la norma koncentriĝo de unu aŭ pluraj aminoacidoj. Se la mankanta substanco estas aldonita, la proteino fariĝos kompleta.
La ĉefaj fontoj de proteinoj de planto kaj besta origino
En la hejma scienca komunumo okupita pri ampleksa studo de manĝa kemio, grupo de profesoroj A.P. Nechaev, liaj kolegoj, kaj studentoj elstaras. La teamo determinis la proteinan enhavon en la ĉefaj manĝaĵoj haveblaj sur la rusa merkato.
- Grava! Identigitaj ciferoj informas pri la proteina enhavo en 100 g de la produkto, liberigita de la manĝebla parto.
- La plej granda kvanto de proteino troviĝas en sojfabo, kukurbaj semoj kaj arakidoj (34,9 - 26,3 g).
- Valoroj de 20 ĝis 30 gramoj troviĝas en semoj, faboj, pistakoj kaj sunfloroj.
- Migdaloj, anacoj, aveloj estas karakterizitaj de nombroj de 15 al 20 gr.
- Nuksoj, pastaĵoj, plej multaj cerealoj (krom rizo, maizo-grajnoj) enhavas de 10 ĝis 15 gramojn da proteino po 100 gramoj da produkto.
- Rizo, maizaj kradoj, pano, ajlo, sekigitaj abrikotoj estas inter 5 kaj 10 gr.
- En 100 gramoj da brasiko, fungoj, terpomoj, prunoj, kelkaj betoj, la enhavo en proteinoj estas de 2 ĝis 5 gramoj.
- Sekvinberoj, rafanoj, karotoj, dolĉaj piproj havas malmultan proteinon, iliaj indikiloj ne superas 2 gramojn.
Se vi ne povis trovi plantan objekton ĉi tie, tiam la proteina koncentriĝo en ĝi estas tro malalta aŭ ĝi tute ne ekzistas. Ekzemple, en fruktaj sukoj estas tre malmulte da proteino, en naturaj vegetalaj oleoj - tute ne.
- La maksimuma proteina koncentriĝo estis trovita en fiŝaj kufoj, malmolaj kaj prilaboritaj fromaĝoj, kaj kuniklo-viando (de 21,1 ĝis 28,9 g).
- Granda nombro da produktoj enhavas de 15 ĝis 10 gramojn da proteino. Ĉi tio estas birdo, mara fiŝo (krom kapelino), brutokarno, salikoko, kalmaroj, dometo, feta fromaĝo, dolĉakva fiŝo.
- Kapelino, kokida ovo, porkaĵo enhavas 12,7 ĝis 15 gramojn da proteino po 100 gramoj da produkto.
- Jogurto, kurba fromaĝo karakterizas per la nombroj 5 - 7,1 gr.
- Lakto, kefir, fermentita bakita lakto, acida kremo, kremo enhavas de 2,8 ĝis 3 gramoj da proteino.
Informoj pri la ĉefaj fontoj de proteinoj de planto kaj besta origino en produktoj, kiuj spertis mult-stadian teknologian prilaboron (kuiraĵon, kolbasojn, ŝinkon, kolbasojn) ne interesas. Ili ne rekomendas por regula sana manĝo. Mallongdaŭra uzo de tiaj produktoj ne gravas.
La rolo de proteino en nutrado
Rezulte de metabolaj procezoj en la korpo, novaj proteinoj-molekuloj konstante formiĝas anstataŭ la malnovaj. La indico de sintezo en malsamaj organoj ne estas la sama. Hormonaj proteinoj, ekzemple, insulino, estas restarigitaj (resintetigitaj) tre rapide, en horoj, minutoj. Proteinoj de la hepato, intestaj mukozoj estas regeneritaj en 10 tagoj. Proteinaj molekuloj de la cerbo, muskoloj, konektiva histo estas restarigitaj la plej longa, restariga sintezo (resintezo) povas daŭri ĝis ses monatoj.
La procezo de uzado kaj sintezo estas karakterizita per nitrogen-ekvilibro.
- En formita homo kun plena sano, la nitrogen-ekvilibro estas nulo. En ĉi tiu kazo, la totala maso de nitrogeno provizita per proteinoj dum nutrado estas egala al la maso eligita el kadukaj produktoj.
- Junaj organismoj rapide disvolviĝas. La bilanco de nitrogeno estas pozitiva. Estas multe da proteino, malpli eltiriĝas.
- En maljuniĝo, malsanuloj, la nitrogen-bilanco estas negativa. La maso de nitrogeno liberigita per metabolaj produktoj estas pli granda ol tiu ricevita per konsumado de nutraĵoj.
La rolo de proteino en nutrado estas provizi al homo la bezonatan kvanton da aminoacidaj komponentoj taŭga por partopreno en la biokemiaj procezoj de la korpo.
Por certigi normalan metabolon, gravas scii kiom da proteino homo bezonas konsumi ĉiutage.
Hejmaj kaj usonaj fiziologoj rekomendas manĝi 0,8 - 1 g da proteino po 1 kg da homa pezo. La nombroj estas sufiĉe averaĝaj. La kvanto dependas de aĝo, naturo de laboro, vivstilo de homo. Averaĝe ili rekomendas konsumadon de 60 gramoj ĝis 100 gramoj da proteino ĉiutage. Por viroj okupiĝantaj pri fizika laboro, la normo povas esti pliigita al 120 gramoj ĉiutage. Por tiuj, kiuj suferas kirurgion, infektajn malsanojn, la normo ankaŭ pliiĝas al 140 gramoj ĉiutage. Diabetaĵoj estas rekomendindaj dietoj kun alta enhavo en proteinaj produktoj, kiuj povas atingi 140g ĉiutage. Homoj kun metabolaj malordoj, inklino al guteto, devas konsumi signife malpli da proteinoj. La normo por ili estas 20 - 40 gramoj tage.
Por homoj okupiĝantaj pri aktivaj sportoj, kiuj pliigas muskolan mason, la normo signife pliiĝas, povas atingi 1,6-1,8 gramojn po 1 kg da atleto.
- Grava! Estas konvene, ke la trejnisto klarigu la respondon al la demando - kiom da proteinoj devas esti konsumataj ĉiutage dum ekzercado. Profesiuloj havas informojn pri energiaj kostoj por ĉiuj specoj de trejnado, manieroj konservi la normalan funkciadon de la korpo de atleto.
Por la efektivigo de ĉiuj fiziologiaj funkcioj gravas ne nur la ĉeesto de esencaj aminoacidoj en la proteino, sed ankaŭ la efikeco de ilia asimilado. Proteinaj molekuloj havas malsamajn nivelojn de organizado, solvebleco, grado de alirebleco al digestaj enzimoj. 96% el laktaj proteinoj, ovoj estas detruitaj efike. En viando, fiŝo, 93-95% de proteinoj estas sekure digestitaj. La escepto estas la proteinoj de la haŭto kaj haroj. Produktoj kun vegetalaj proteinoj estas digestitaj de 60-80%. En legomoj, 80% de proteinoj estas sorbitaj, en terpomoj - 70%, en pano - 62-86%.
La rekomendinda porcio de proteinoj el bestaj fontoj devas esti 55% de la tuta kvanto de proteina maso.
- Proteina manko en la korpo kondukas al signifaj metabolaj ŝanĝoj. Tiaj patologioj nomiĝas distrofio, kwashiorkor. Por la unua fojo, malobservo estis rivelita ĉe la loĝantoj de la sovaĝaj triboj de Afriko, karakterizita de negativa bilanco de nitrogeno, difektita intesta funkcio, muskola atrofio kaj senĉesa kresko. Parta proteina manko povas okazi kun similaj simptomoj, kiuj povas esti mildaj dum iom da tempo. Precipe danĝera estas la manko de proteino en la korpo de la infano. Tiaj dietaj malordoj povas provoki la fizikan kaj intelektan malsuperecon de kreskanta homo.
- Troa proteino en la korpo troŝarĝas la ekskretan sistemon. La ŝarĝo sur la renoj pliiĝas. Kun ekzistantaj patologioj en la rena histo, la procezo povas pligravigi. Estas tre malbone, se troo de proteino en la korpo estas akompanata de manko de aliaj valoraj manĝaĵoj. En antikvaj tempoj, en la landoj de Azio ekzistis metodo de ekzekuto, per kiu la kondamnito estis manĝita nur viando. Rezulte la kriminto mortis pro la formado de putraj produktoj en la intesto, post ĉi tiu veneniĝo.
Akceptebla aliro al provizado de la korpo kun proteino garantias efikan funkciadon de ĉiuj vivsistemoj.
Studu historion

La proteino estis unuafoje akirita (en formo de gluten) en 1728 de la italo Jacopo Bartolomeo Beccari el tritika faruno. Proteinoj estis izolitaj en apartan klason de biologiaj molekuloj en la 18-a jarcento kiel rezulto de la laboro de la franca kemiisto Antoine de Fourcroix kaj aliaj sciencistoj, kiuj rimarkis la posedaĵon de proteinoj koaguli (denature) sub la influo de varmego aŭ acidoj. En tiu tempo, proteinoj kiel albumino ("ovo-blanka"), fibrino (proteino el la sango) kaj gluteno el tritikaj grajnoj estis esploritaj.
Komence de la 19a jarcento, oni jam informis pri la elementa konsisto de proteinoj; oni sciis, ke aminoacidoj formiĝas dum hidrolizo de proteinoj. Iuj el ĉi tiuj aminoacidoj (ekz. Glicino kaj leŭkino) jam karakteriziĝis. Surbaze de la analizo de la kemia konsisto de proteinoj, la nederlanda kemiisto Gerrit Mulder hipotezis, ke preskaŭ ĉiuj proteinoj havas similan empirian formulon. En 1836, Mulder proponis la unuan modelon de la kemia strukturo de proteinoj. Surbaze de la teorio de radikaloj, post pluraj rafinadoj li alvenis al la konkludo, ke la minimuma struktura unuo de proteino havas la jenan kunmetaĵon: C40H62N10Ho12. Li nomis ĉi tiun unuon "proteino" (Pr) (el la greka. Protos - la unua, primara), kaj la teorion - "proteina teorio". La esprimo "proteino" mem estis proponita de la sveda kemiisto Jacob Berzelius. Laŭ Mulder, ĉiu proteino konsistas el pluraj protein-unuoj, sulfuro kaj fosforo. Ekzemple, li sugestis skribi la fibrinan formulon kiel 10PrSP. Mulder ankaŭ esploris la produktojn de detruo de proteinoj - aminoacidoj kaj por unu el ili (leŭcino) kun malgranda frakcio de eraro, li determinis la molekulan pezon - 131 daltons. Kun la amasiĝo de novaj datumoj pri proteinoj, la teorio pri proteino komencis esti kritikata, sed malgraŭ tio, ĝis la malfruaj 1850-aj jaroj ĝi estis ankoraŭ konsiderata universale rekonita.
Antaŭ la fino de la 19-a jarcento, la plej multaj el la aminoacidoj kiuj konsistigas proteinojn estis esploritaj. En la malfruaj 1880-aj jaroj. Rusa sciencisto A. Ya.Danilevskij rimarkis la ekziston de peptidaj grupoj (CO - NH) en la proteina molekulo. En 1894 la germana fiziologo Albrecht Kossel proponis teorion laŭ kiu aminoacidoj estas la ĉefaj strukturaj elementoj de proteinoj. Komence de la 20a jarcento, la germana kemiisto Emil Fischer pruvis eksperimente, ke proteinoj konsistas el aminoacidaj restaĵoj ligitaj per peptidaj ligoj. Li ankaŭ efektivigis la unuan analizon de la aminoacida sinsekvo de la proteino kaj klarigis la fenomenon de proteolizo.
Tamen la centra rolo de proteinoj en organismoj ne agnoskis ĝis 1926, kiam la usona kemiisto James Sumner (poste Nobel-premio pri kemio) montris, ke la ureasa enzimo estas proteino.
La malfacileco en izolado de puraj proteinoj malfaciligis studadon. Tial la unuaj studoj estis farataj uzante tiujn polipeptidojn, kiuj facile povus esti purigitaj en grandaj kvantoj, tio estas, sangaj proteinoj, kokaj ovoj, diversaj toksinoj, kaj ankaŭ digestajn / metabolajn enzimojn kaŝitajn post buĉado de brutoj. En la malfruaj 1950-aj jaroj, la kompanio Armor Hot Dog Co. sukcesis demeti kilogramon da bovina pankreata ribonuklease A, kiu fariĝis eksperimenta objekto por multaj studoj.
La ideo, ke la malĉefa strukturo de proteinoj estas la rezulto de la formado de hidrogenaj ligoj inter restaĵoj de aminoacidoj estis proponita de William Astbury en 1933, sed Linus Pauling estas konsiderata la unua sciencisto, kiu sukcesis sukcese antaŭdiri la malĉefan strukturon de proteinoj. Poste, Walter Kausman, surbaze de la laboro de Kai Linnerstrom-Lang, faris gravan kontribuon por kompreni la leĝojn de la formado de la tercia strukturo de proteinoj kaj la rolon de hidrofobaj interagoj en ĉi tiu procezo. En la malfruaj 1940-aj jaroj kaj fruaj 1950-aj jaroj, Frederick Senger disvolvis metodon por sekvencado de proteinoj, per kiu li determinis la aminoacidan sekvencon de du insulinĉenoj antaŭ 1955, pruvante ke proteinoj estas linearaj polimeroj de aminoacidoj, kaj ne branĉitaj (kiel iuj sukeroj. ) ĉenoj, koloidoj aŭ cikloloj. La unua proteino, kies sinsekvo de aminoacidoj estis establita de sovetiaj / rusaj sciencistoj, estis en 1972 aspartato aminotransferase.
La unuaj spacaj strukturoj de proteinoj akiritaj per difraktado de radioj X (analizo de difraktado de radioj X) konatiĝis fine de la 1950-aj jaroj kaj fruaj 1960-aj jaroj, kaj strukturoj malkovritaj uzante nuklean magnetan resonancon en la 1980-aj jaroj. En 2012, la Datenbanko pri Proteinoj enhavis proksimume 87.000 proteinajn strukturojn.
En la 21a jarcento, la esplorado pri proteinoj translokiĝis al kvalite nova nivelo, kiam oni studas ne nur unuopajn purigitajn proteinojn, sed ankaŭ la samtempan ŝanĝon de la nombro kaj post-tradukaj modifoj de granda nombro da proteinoj de unuopaj ĉeloj, histoj aŭ tutaj organismoj. Ĉi tiu areo de biokemio nomiĝas proteomiko. Uzante bioinformatikajn metodojn, fariĝis eble ne nur prilabori datumajn analizajn difraktajn radiografiojn, sed ankaŭ antaŭdiri la strukturon de la proteino surbaze de ĝia aminoacida sinsekvo. Nuntempe krioelektronika mikroskopio de grandaj proteinaj kompleksoj kaj la prognozo de la spacaj strukturoj de proteinaj domajnoj uzantaj komputilajn programojn alproksimiĝas al atoma precizeco.

Proteina grandeco povas esti mezurita koncerne aminoacidajn restaĵojn aŭ en daltonoj (molekula pezo), sed pro la relative granda grandeco de la molekulo, la proteina maso estas esprimita en derivitaj unuoj - kilodaltonoj (kDa). Legomaj proteinoj, averaĝe, konsistas el 466 aminoacidaj restaĵoj kaj havas molekulan pezon de 53 kDa. La plej granda proteino nuntempe konata - titino - estas ero de muskolaj sarcomeres, la molekula pezo de siaj diversaj variantoj (izoformoj) varias en la gamo de 3000 ĝis 3700 kDa. Titino de la soleus muskolo (lat. Soleus) de homo konsistas el 38.138 aminoacidoj.
Amfoterio
Proteinoj havas la posedaĵon de amfoterio, tio estas, laŭ la kondiĉoj, ili montras ambaŭ acidajn kaj bazajn propraĵojn. En proteinoj, ekzistas pluraj specoj de kemiaj grupoj kapablaj ionigi en akva solvo: karboxilaj restaĵoj de la flankaj ĉenoj de acidaj aminoacidoj (aspartaj kaj glutamaj acidoj) kaj nitrogenaj enhavantaj grupoj de la flankaj ĉenoj de bazaj aminoacidoj (ĉefe la ε-amino grupo de lizino kaj la amidina restaĵo CNH (NH)2) arginino, en iomete malpli granda - la restaĵo de imidazola histidino). Ĉiu proteino estas karakterizita per izoelektra punkto (pI) - meza acideco (pH), ĉe kiu la tuta elektra ŝarĝo de la molekuloj de ĉi tiu proteino estas nulo kaj sekve ili ne moviĝas en la elektra kampo (ekzemple per elektroforesis). Ĉe izoelektra punkto, hidratado kaj solvebleco de la proteino estas minimumaj. La pI-valoro dependas de la proporcio de acidaj kaj bazaj aminoacidaj restaĵoj en proteino: por proteinoj enhavantaj multajn acidajn aminoacidajn restaĵojn, izoelektraj punktoj kuŝas en la acida regiono (tiaj proteinoj nomiĝas acidaj), kaj ĉe proteinoj enhavantaj pli bazajn restaĵojn, ili estas en la alkala regiono (bazaj proteinoj) ) La pI-valoro de ĉi tiu proteino ankaŭ povas varii depende de la ionika forto kaj la speco de buĉa solvo en kiu ĝi troviĝas, ĉar neŭtralaj saloj influas la gradon de jonigo de la kemiaj grupoj de la proteino. La pI de proteino povas esti determinita, ekzemple, de titoliga kurbo aŭ per izoelektra fokuso.
Ĝenerale, la pI de proteino dependas de la funkcio, kiun ĝi plenumas: la izoelektra punkto de plej multaj proteinoj en vertebrulaj histoj iras de 5,5 ĝis 7,0, sed en iuj kazoj la valoroj kuŝas en ekstremaj areoj: ekzemple, por pepsino, proteolitika enzimo de forte acida gastriko suko pI
1, kaj por salminoj - la protamina proteino de salma lakto, kies ĉefaĵo estas alta arginina enhavo - pI
12. Proteinoj, kiuj ligas al nukleaj acidoj pro elektrostatika interagado kun fosfataj grupoj ofte estas la ĉefaj proteinoj. Ekzemplo de tiaj proteinoj estas histonoj kaj protaminoj.
Kio estas proteinoj?
Proteinoj estas altaj molekulaj pezaj kompleksaj organikaj komponaĵoj konsistantaj el restaĵoj de aminoacidoj, kombinitaj en speciala maniero. Ĉiu proteino havas sian propran individuan aminoacidan sekvencon, sian lokon en la spaco. Gravas kompreni, ke la proteinoj enirantaj en la korpon ne estas sorbitaj de ili senŝanĝe, ili estas dividitaj en aminoacidojn kaj kun ilia helpo la korpo sintezas siajn proteinojn.
22 aminoacidoj partoprenas en la formado de proteinoj, 13 el ili povas esti transformitaj unu al la alia, 9 - fenilalanino, triptofano, lizino, histidino, treonino, leŭkino, valino, izoleucino, metionino - estas neanstataŭigeblaj. La manko de neŝanĝeblaj acidoj en la korpo estas neakceptebla, ĉi tio kondukos al interrompo de la korpo.
Gravas ne nur la fakto, ke la proteino eniras la korpon, sed ankaŭ el kiuj aminoacidoj ĝi konsistas!

Kio estas proteino?
Proteinoj (proteinoj / polipeptidoj) - organikaj substancoj, naturaj polimeroj enhavantaj dudek rilatajn aminoacidojn. Kombinoj provizas multajn specojn. La korpo trafas la sintezon de dek du interŝanĝeblaj aminoacidoj mem.
Ok el la dudek esencaj aminoacidoj trovitaj en proteino ne povas sintezi la korpon memstare, ili estas akiritaj per manĝaĵo. Ĉi tiuj estas valino, leŭkino, izoleŭkino, metionino, triptofano, lizino, treonino, fenilalanino, kiuj gravas por la vivo.
Kio okazas proteino
Distingu inter besto kaj vegetaĵo (laŭ deveno). Du specoj de konsumo estas bezonataj.
Besto:
Ovo blanka estas facile kaj preskaŭ komplete sorbata de la korpo (90-92%). Proteinoj de fermentitaj laktaĵoj estas iomete pli malbonaj (ĝis 90%). Proteinoj de freŝa tuta lakto estas absorbitaj eĉ malpli (ĝis 80%).
La valoro de bovaĵo kaj fiŝo en la plej bona kombinaĵo de esencaj aminoacidoj.
Legomo:
Sojko, kanolo kaj kukolo havas bonan aminoacidproporcion por la korpo. Ĉe kultivaĵoj, tiu rilatumo estas pli malforta.
Ne ekzistas produkto kun ideala aminoacida proporcio. Ĝusta nutrado implikas kombinaĵon de bestaj kaj vegetalaj proteinoj.
La bazo de nutrado "laŭ la reguloj" estas besta proteino. Ĝi estas riĉa je esencaj aminoacidoj, kaj donas bonan absorbadon de vegeta proteino.
Proteinoj funkcias en la korpo
En la ĉeloj de la histo, ĝi plenumas multajn funkciojn:
- Protekta. La funkciado de la imunsistemo estas neŭtraligo de fremdaj substancoj. Produktado de antikorpoj.
- Transportado. Provizado de diversaj substancoj, ekzemple hemoglobino (provizo de oksigeno).
- Regule. Subteni hormonal fonon.
- Propulso. Ĉiuj specoj de movado provizas actinon kaj miozinon.
- Plasta. La kondiĉo de la konektebla histo estas kontrolita de la kolagena enhavo.
- Kataluna. Ĝi estas katalizilo kaj akcelas la paŝon de ĉiuj biokemiaj reagoj.
- Konservado kaj transdono de genaj informoj (molekuloj de DNA kaj RNA).
- Energio. Provizado de la tuta korpo per energio.
Aliaj provizas spiradon, respondecas pri la digesto de manĝaĵoj, reguligas metabolon. La fotosensema rodopsina proteino respondecas pri vida funkcio.
Sangaj glasoj enhavas elastinon, danke al ĝi ili plene funkcias. La fibrinogena proteino provizas sangan koaguladon.
Simptomoj de manko de proteino en la korpo
Proteina manko estas sufiĉe ofta okazo kun subnutrado kaj hiperaktiva vivstilo de moderna homo. Milda formo ĝi estas esprimata en regula laceco kaj malbona agado. Kun kresko de nesufiĉaj kvantoj, la korpo signalas per simptomoj:
- Ĝenerala malforteco kaj kapturno. Malpliiĝis humoro kaj aktiveco, apero de muskola laceco sen speciala fizika streĉo, malgrava kunordigo de movadoj, malfortiĝo de atento kaj memoro.
- Kapdoloroj kaj plimalboniga dormo. La rezulta sendormeco kaj angoro indikas mankon de serotonino.
- Ofte murmuroj, gruntoj. Manko de enzimoj kaj hormonoj provokas elĉerpiĝon de la nerva sistemo: irritabilidad pro iu ajn kialo, nepravigebla agresemo, emocia bremsado.
- Pale haŭto, erupcioj. Kun manko de fera enhavanta proteino, anemio disvolviĝas, kies simptomoj estas sekeco kaj paleco de la haŭto, mukozaj membranoj.
- Ŝvelado de la ekstremaĵoj. Malalta plasmo-proteino enhavas la akvon-salan ekvilibron. Subkutana graso akumulas fluidon en la maleoloj.
- Malbona resanigo de vundoj kaj abrazioj. Ĉela riparo estas malhelpita pro manko de "konstrua materialo".
- Malfortikeco kaj hara perdo, malforteco de najloj. La apero de forvelko pro seka haŭto, exfoliado kaj krakado de la najla plato estas la plej ofta signalo de la korpo pri manko de proteino. Haro kaj ungoj konstante kreskas kaj senprokraste respondas al manko de substancoj, kiuj antaŭenigas kreskon kaj bonan staton.
- Malzorgema pezoperdo. La malapero de kilogramoj pro neniu ŝajna kialo estas pro la bezono de la korpo kompensi la mankon de proteino pro muskola maso.
- Malsukceso de la koro kaj sangaj glasoj, apero de manko de spiro. La spiraj, digestaj kaj genitourinaj sistemoj ankaŭ plimalboniĝas. Disnea aperas sen fizika streĉo, tuso sen malvarmoj kaj viraj malsanoj.
Kun apero de simptomoj de ĉi tiu speco, vi devas tuj ŝanĝi la reĝimon kaj kvaliton de manĝaĵo, rekonsideri vian vivmanieron, kaj se pligravigi, konsulti kuraciston.
Kiom da proteino bezonas por asimilado
La konsumokosto ĉiutage dependas de aĝo, sekso, speco de laboro. Datumoj pri la normoj estas prezentitaj en la tabelo (sube) kaj estas desegnitaj por normala pezo.
Disbatanta proteinan konsumon plurfoje estas nedeviga. Ĉiu determinas konvenan formon por si mem, la ĉefa afero estas konservi la ĉiutagan konsuman indicon.
| Laborista agado + |
fizika aktiveco
Alta proteina enhavo en nutraĵoj
Agnoskitaj manĝaĵoj kun proteinoj:
El ĉiuj varioj de viando, la unua loko post kortega enhavo estos bovaĵo: 18,9 g. Post ĝi porkaĵo: 16,4 g, ŝafido: 16,2 g.
Mariskoj kaj kalmaroj estas la ĉefoj: 18,0 g.
La plej riĉa fiŝo por proteino estas salmo: 21,8 g, tiam rozkolora salmo: 21 g, pika poĉo: 19 g, makro: 18 g, haringo: 17,6 g kaj moruo: 17,5 g.
Inter laktaĵoj, kefir kaj acida kremo firme tenas la pozicion: 3,0 g, poste lakto: 2,8 g.
Altaj Grajnoj - Hércules: 13,1 g, mijo: 11,5 g, semolo: 11,3 g
Sciante la normon kaj konsiderante financajn ŝancojn, vi povas ĝuste formi menuon kaj nepre kompletigi ĝin per grasoj kaj karbonhidratoj.
La rilatumo de proteino en nutrado
La proporcio de proteinoj, grasoj, karbonhidratoj en sana dieto devas esti (en gramoj) 1: 1: 4. La ŝlosilo de la ekvilibro de sana plado povas esti reprezentita alimaniere: proteinoj 25-35%, grasoj 25-35%, karbonhidratoj 30-50%.
Samtempe grasoj devas esti utilaj: oliva aŭ lina oleo, nuksoj, fiŝoj, fromaĝo.
Karbonhidratoj en telero estas malmolaj pastoj, ajnaj freŝaj legomoj, same kiel fruktoj / sekigitaj fruktoj, acida laktaĵoj.
Proteinoj laŭ porcioj elekteblas kombinitaj: vegetaĵoj + bestoj.
Aminoacidoj Enhavitaj En Proteinoj
La interŝanĝebleco povas sintezi la korpon mem, sed ilia provizo el la ekstero neniam estas superflua. Precipe kun aktiva vivstilo kaj granda fizika praktikado.
Ĉiuj senescepte gravas, la plej popularaj el ili:
Alanine.
Ĝi stimulas la metabolon de karbonhidratoj, helpas forigi toksinojn. Respondeca pri "pureco". Alta enhavo en viando, fiŝo, laktaĵoj.
Arginino.
Necesas kontrakti muskolojn, sanan haŭton, kartilagojn kaj artikojn. Provizas grasan bruladon kaj imunsistemon. Ĝi estas en ĉia viando, lakto, ajnaj nuksoj, gelateno.
Aspartika acido.
Provizas energian ekvilibron. Plibonigas la funkciecon de la centra nerva sistemo. Nu replenigu la energian rimedon de bovaĵoj kaj kokaj pladoj, lakto, kano sukero. Enhavita en terpomoj, nuksoj, cerealoj.
Histidino.
La ĉefa "konstruanto" de la korpo transformiĝas en histaminon kaj hemoglobinon. Rapide resanigas vundojn, respondecas pri kreskaj mekanismoj. Relative multe en lakto, cerealoj kaj ia ajn viando.
Serena.
Neŭrotransmisilo, nemalhavebla por la klara funkciado de la cerbo kaj centra nerva sistemo. Estas arakidoj, viando, cerealoj, sojfabo.
Kun taŭga nutrado kaj ĝusta vivmaniero, ĉiuj aminoacidoj aperos en la korpo por sintezo de "kuboj" kaj modeligado de sano, beleco kaj longeveco.
Kio kaŭzas mankon de proteino en la korpo
- Ofte infektaj malsanoj, malfortiĝo de la imunsistemo.
- Streso kaj angoro.
- Maljuniĝo kaj malrapidigo de ĉiuj metabolaj procezoj.
- Flanka efiko de la uzo de unuopaj medikamentoj.
- Malsukcesoj en la digesta vojo.
- Vundoj.
- Manĝaĵo bazita sur rapida manĝo, tujaj produktoj, duonfinitaj produktoj de malalta kvalito.
Manko de ununura aminoacido ĉesigos la produktadon de specifa proteino. La korpo estas organizita laŭ la principo "plenigi la malplenojn", do la mankantaj aminoacidoj estos ĉerpitaj el aliaj proteinoj. Ĉi tiu "rekonstruado" malhelpas la funkciadon de organoj, muskoloj, koro, cerbo kaj poste estigas la malsanon.
Proteina manko en infanoj malhelpas kreskon, kaŭzas fizikajn kaj mensajn handikapojn.
La disvolviĝo de anemio, la apero de haŭtaj malsanoj, patologio de osto kaj muskola histo ne estas kompleta listo de malsanoj. Severa proteina distrofio povas rezultigi malsaniĝon kaj kvashiorkor (speco de severa distrofio pro manko de proteinoj).
Kiam proteino malbonigas la korpon
- troa ricevo
- kronikaj malsanoj de la hepato, renoj, koro kaj sangaj glasoj.
Supertroprovizo ne ofte okazas pro nekompleta absorbo de substanco fare de la korpo.Ĝi okazas ĉe tiuj, kiuj volas pliigi muskolon kiel eble plej rapide sen sekvi la rekomendojn de trejnistoj kaj nutristiistoj.
La problemoj pri "troa" ricevo inkluzivas:
Reala fiasko. Troaj kvantoj de proteinoj superŝarĝas organojn, interrompante sian naturan funkcion. "Filtrilo" ne povas trakti la ŝarĝon, rena malsano aperas.
Hepato-malsano. Troa proteino amasigas amoniakon en la sango, kiu plimalbonigas la kondiĉon de la hepato.
La disvolviĝo de aterosklerozo. Plej multaj bestaj produktoj, krom utilaj substancoj, enhavas malutilajn grasojn kaj kolesterolon.
Homoj, kiuj suferas patologion de la hepato, renoj, kardiovaskulaj kaj digestaj sistemoj, devas limigi la konsumon de proteinoj.
Prizorgo pri propra sano estas rekompencita bonvole al tiuj, kiuj zorgas pri ĝi. Por eviti severajn konsekvencojn, vi devas memori la bezonon de la korpo por resaniĝo. Plena ripozo, nutrado, vizitantaj specialistoj plilongigos junecon, sanon kaj vivon.
Solubileco
Proteinoj varias je grado de soleco en akvo. Akvo-solveblaj proteinoj estas nomataj albumino, ĉi tiuj inkluzivas sangajn kaj laktajn proteinojn. Inter nesolveblaj aŭ skleroproteinoj inkluzivas, ekzemple, keratino (la proteino kiu konsistigas harojn, harojn de mamuloj, plumojn de birdoj, ktp.) Kaj fibroinon, kiu estas parto de silko kaj teksaĵo. La solvebleco de proteino estas determinita ne nur de ĝia strukturo, sed de eksteraj faktoroj, kiel la naturo de la solvilo, jona forto, kaj la pH de la solvo.
Proteinoj estas ankaŭ dividitaj en hidrofilajn (akvorezistajn) kaj hidrofobajn (akvoregentajn). Plej multaj proteinoj de la citoplasmo, kerno kaj intercelula substanco, inkluzive de nesolveblaj keratino kaj fibroino, estas hidrofilaj. Plej multaj proteinoj, kiuj konsistigas biologiajn membranojn, estas hidrofobaj - integraj membranaj proteinoj, kiuj interagas kun hidrofobaj lipidoj de la membrano (ĉi tiuj proteinoj, kiel regulo, havas ankaŭ hidrofilajn lokojn).
Biosintezo de proteinoj en la korpo
Biosintezo de proteinoj - la formado en la korpo de la dezirataj proteinoj el aminoacidoj kombinante ilin kun speciala speco de kemia ligo - la polipeptida ĉeno. DNA stokas informojn pri proteina strukturo. La sintezo mem okazas en speciala parto de la ĉelo nomata ribosoma. RNA trapasas informojn de la dezirata geno (DNA-loko) al la ribosomo.
Pro tio ke proteina biosintezo estas multfaceta, kompleksa, uzas la informojn bazitajn sur la homa ekzisto - DNA, ĝia kemia sintezo estas malfacila tasko. Sciencistoj lernis kiel akiri inhibilojn de iuj enzimoj kaj hormonoj, sed la plej grava scienca tasko estas akiri proteinojn per genetika inĝenierado.

Transportado
La transporta funkcio de speciala sanga proteino - hemoglobino. Danke al ĉi tiu proteino, oksigeno liveras de la pulmoj al la organoj kaj histoj de la korpo.
Ĝi konsistas en la agado de proteinoj de la imunsistemo nomataj antikorpoj. Temas pri antikorpoj, kiuj gardas la sanon de la korpo, protektante ĝin kontraŭ bakterioj, virusoj, venenoj, kaj permesas sangon formi kozon anstataŭ malferma vundo.
La funkcia signalo de proteinoj estas transdoni signalojn (informojn) inter ĉeloj.
Normaj Proteinoj por Plenkreskulo
La bezono de la homa korpo por proteino rekte dependas de ĝia fizika aktiveco. Ju pli ni moviĝas, des pli rapide ĉiuj biokemiaj reagoj okazas en nia korpo. Homoj, kiuj regule faras ekzercadon, bezonas preskaŭ duoble pli da proteino ol la averaĝa homo. Manko de proteino por homoj en sportoj estas danĝera "sekigi" la muskolojn kaj elĉerpiĝon de la tuta korpo!
Averaĝe, la proteina normo por plenkreskulo estas kalkulata surbaze de koeficiento de 1 g da proteino po 1 kg da pezo, tio estas proksimume 80-100 g por viroj, 55-60 g por virinoj. Viraj atletoj konsilas pliigi la kvanton de proteino konsumita al 170-200 g ĉiutage.

Taŭga proteina nutrado por la korpo
Taŭga nutrado por saturi la korpon per proteino estas kombinaĵo de bestaj kaj plantaj proteinoj. La grado de asimilado de proteino el manĝaĵo dependas de ĝia deveno kaj la metodo de varmotraktado.
Tiel ĉirkaŭ 80% de la tuta konsumado de besta proteino kaj 60% de vegeta proteino estas sorbita de la korpo. Produktoj de besta origino enhavas pli grandan kvanton da proteino por unueca maso de la produkto ol en vegetaĵoj. Krome, la kunmetaĵo de "bestaj" produktoj inkluzivas ĉiujn aminoacidojn, kaj plantaj produktoj tiurilate estas konsideritaj malsuperaj.
Bazaj nutraj reguloj por pli bona absorción de proteinoj:
- Milda maniero kuiri - kuiri, vapori, kuvi. Fritado devas esti forĵetita.
- Oni rekomendas manĝi pli da fiŝoj kaj birdoj. Se vi vere volas viandon - elektu bovaĵon.
- Buljonoj devas esti ekskluditaj de la dieto, ili estas grasaj kaj malutilaj. En ekstremaj kazoj, vi povas kuiri la unuan pladon per la "malĉefa buljono".
Trajtoj de proteina nutrado por muskola kresko
Atletoj, kiuj aktive gajnas muskolan mason, devas aliĝi al ĉiuj supraj rekomendoj. Plejparto de ilia dieto devas esti proteinoj de besta origino. Ili devas esti manĝitaj lige kun vegetalaj proteinaj produktoj, el kiuj sojfabo estas aparta prefero.
Ankaŭ necesas konsulti kuraciston kaj pripensi la uzon de specialaj proteinaj trinkaĵoj, kies procento de proteino absorbas 97–98%. La specialisto individue elektos trinkaĵon, kalkulos la ĝustan dozon. Ĉi tio estos agrabla kaj utila proteina suplemento al fortika trejnado.
Denaturado
Proteino-denaturado rilatas al iuj ajn ŝanĝoj en ĝia biologia aktiveco kaj / aŭ fiziokemiaj proprietoj asociitaj kun la perdo de kvaternara, terciara aŭ sekundara strukturo (vidu la sekcion "strukturo de proteinoj"). Kiel regulo, proteinoj estas sufiĉe stabilaj sub tiuj kondiĉoj (temperaturo, pH, ktp), en kiuj ili normale funkcias en la korpo. Akuta ŝanĝo en ĉi tiuj kondiĉoj kondukas al proteino-denaturado. Depende de la naturo de la denaturiga agento, mekanika (forta agito aŭ skuado), fizika (hejtado, malvarmigo, radiado, sonado) kaj kemiaj (acidaj kaj alkalis, surfactants, ureo) distingiĝas.
La denaturado de proteinoj povas esti kompleta aŭ parta, revertebla aŭ neinversigebla. La plej fama kazo de neinversigebla proteino-denaturado en ĉiutaga vivo estas preparado de koka ovo, kiam, sub influo de alta temperaturo, la akvorezista travidebla proteino ovalbumino fariĝas densa, nesolvebla kaj opaka. Denaturado en iuj kazoj estas revertebla, kiel en la kazo de precipitaĵo de akvo-solveblaj proteinoj uzantaj salojn de amonio (metodo de salado), kaj ĉi tiu metodo estas uzata kiel maniero purigi ilin.
Proteinaj molekuloj estas linearaj polimeroj konsistantaj el restaĵoj de α-L-aminoacidoj (kiuj estas monomeroj), kaj ankaŭ modifitaj restaĵoj de aminoacidoj kaj komponantoj de ne-aminoaciaj naturoj povas esti inkluzivitaj en la kunmetaĵo de proteinoj. En la scienca literaturo, unu- aŭ tri-literaj mallongigoj estas uzataj por rilati aminajn acidojn. Kvankam al unua vido povas ŝajni, ke la uzo de "nur" 20 specoj de aminoacidoj en plej multaj proteinoj limigas la diversecon de proteinaj strukturoj, fakte la nombro de ebloj malfacile povas esti supraestimata: por ĉeno de 5 aminoacidaj restaĵoj, ĝi jam superas 3 milionojn, kaj ĉeno de 100 aminoacidaj restaĵoj. (malgranda proteino) povas esti reprezentita en pli ol 10.130 variantoj. Proteinoj de 2 ĝis kelkaj dekoj da restaĵoj de aminoacidoj longe estas ofte nomataj peptidoj, kun pli granda grado de polimerigo - sciurojkvankam ĉi tiu divido estas tre arbitra.
Kiam proteino formiĝas kiel rezulto de la interagado de α-karboxila grupo (-COOH) de unu aminoacido kun α-amino-grupo (-NH)2) de alia aminoacido, peptidaj ligoj formiĝas. La finoj de la proteino nomiĝas la finaĵo N- kaj C-depende de kiu el la grupoj de la fina aminoacida restaĵo estas libera: -NH2 aŭ -COOH respektive. En proteina sintezo sur la ribosoma, la unua (N-fina stacio) aminoacida restaĵo estas kutime la metionina restaĵo, kaj postaj restaĵoj estas ligitaj al la C-finaĵo de la antaŭa.
Trajtoj de proteina nutrado, dieters
Tiuj, kiuj volas perdi pezon, devas manĝi bestojn kaj vegetalajn proteinajn produktojn. Gravas apartigi ilian konsumon, ĉar la tempo por ilia asimilado estas malsama. Grasaj viandaj produktoj devas esti forĵetitaj, oni ne devas misuzi terpomojn, oni devas preferi cerealojn kun averaĝa proteino.
Ne iru al ekstremoj kaj "sidiĝu" sur proteina dieto. Ĝi ne plaĉas al ĉiuj, ĉar la kompleta ekskludo de karbonhidratoj kondukos al malpliigo de laboranta kapablo kaj energio. Sufiĉas manĝi matenojn kun karbonhidratoj matene - tio donos energion dum la tago, posttagmeze manĝu malaltan proteinan proteinon. Por kompensi la mankon de energio vespere, la korpo komencos bruligi korpan grason, tamen ĉi tiu procezo estos sekura por la sano de la korpo.
Nepre inkluzivas ĝustajn kaj taŭge pretajn proteinajn manĝaĵojn en via dieto. Por la korpo, proteino estas la ĉefa konstrua materialo! Kune kun regula trejnado, ĝi helpos vin konstrui belan atletan korpon!
Proteinoj estas la plej gravaj kemiaj komponaĵoj, sen kiuj la esenca aktiveco de la korpo estus neebla. Proteinoj konsistas el enzimoj, ĉeloj de organoj, histoj. Ili respondecas pri metabolaj, transportaj kaj multaj aliaj procezoj okazantaj en la homa korpo. Proteinoj ne povas akumuliĝi "en rezervo", tial ili devas esti regule ingestitaj. Ili aparte gravas por homoj, kiuj okupiĝas pri sportoj, ĉar proteinoj estas reguligitaj.
Organizaj Niveloj

K. Lindstrom-Lang proponis distingi 4 nivelojn de la struktura organizo de proteinoj: primara, malĉefa, terciara kaj kvaternara strukturoj. Kvankam ĉi tiu divido estas iom malaktuala, ĝi daŭre uziĝas. La primara strukturo (vico de restaĵoj de aminoacidoj) de polipeptido estas determinita de la strukturo de sia geno kaj genetika kodo, kaj strukturoj de pli altaj ordoj formiĝas dum faldado de proteinoj. Kvankam la spaca strukturo de la proteino entute estas determinita de ĝia aminoacida sinsekvo, ĝi estas sufiĉe laborema kaj povas dependi de eksteraj kondiĉoj, tial pli ĝustas paroli pri la preferata aŭ plej energie favora proteino.
Primara strukturo

La primara strukturo estas la vico de restaĵoj de aminoacidoj en la polipeptida ĉeno. La primara strukturo de proteino estas tipe priskribita uzante nomumojn ununurajn aŭ tri por restaĵoj de aminoacido.
Gravaj ecoj de la primara strukturo estas konservativaj motivoj - stabilaj kombinaĵoj de aminoacidaj restaĵoj, kiuj plenumas certan funkcion kaj troviĝas en multaj proteinoj. Konservativaj motivoj estas konservataj dum la evoluo de specioj; ofte eblas antaŭdiri la funkcion de nekonata proteino el ili. La grado de homologio (simileco) de la aminoacidaj sekvencoj de proteinoj de malsamaj organismoj povas esti uzata por taksi la evoluan distancon inter la taksonoj al kiuj apartenas ĉi tiuj organismoj.
La primara strukturo de proteino povas esti determinita per sekvencaj metodoj de proteinoj aŭ per la primara strukturo de ĝia ARNm per la genetika kodotablo.
Malĉefa strukturo
La malĉefa strukturo estas la loka ordigo de fragmento de polipeptida ĉeno stabiligita de hidrogenaj ligoj.Jen la plej oftaj specoj de sekundara proteina strukturo:
- α-helicoj estas densaj turnoj ĉirkaŭ la longa akso de la molekulo. Unu turno estas 3,6 aminoacidaj restaĵoj, la helico-tonalto estas 0,54 nm (0,15 nm falas sur unu aminoacida restaĵo). La spiralo stabiligas per hidrogenaj ligoj inter la H kaj O-peptidaj grupoj, interspacigitaj 4 unuojn dise. Kvankam la α-helico povas esti aŭ maldekstramana aŭ dekstramana, dekstraj superregas en proteinoj. La spiralo estas malatentigita de la elektrostataj interagoj de glutamika acido, lizino, arginino. Proksime unu al la alia, asparagino, serino, treonino kaj leŭkina restaĵoj povas sterike intermiksiĝi en la formadon de la helico, prolinaj restaĵoj kaŭzas katenon kaj ankaŭ detruas α-helicojn,
- β-tukoj (falditaj tavoloj) estas pluraj zigzagaj polipeptidaj ĉenoj en kiuj hidrogenaj ligoj formiĝas inter relative malproksimaj aminoacidoj (0,34 nm per aminoacida restaĵo) en la primara strukturo aŭ malsamaj proteinaj ĉenoj (anstataŭ ol proksime interspacigitaj, kiel estas la kazo. esti en la α-helico). Ĉi tiuj ĉenoj kutime estas direktitaj per la N-finoj en kontraŭaj direktoj (kontraŭparalela orientiĝo) aŭ en unu direkto (paralela β-strukturo). Eblas ankaŭ la ekzisto de miksita β-strukturo konsistanta el paralelaj kaj kontraŭparalelaj β-strukturoj. Por formado de β-litoj gravas malgrandaj grandecoj de flankaj grupoj de aminoacidoj, kutime superregas glicino kaj alanino,
- π-helico
- 310spiraloj
- malordigitaj fragmentoj.
Terciara strukturo

La terciara strukturo estas la spaca strukturo de la polipeptida ĉeno. Strukture, ĝi konsistas el elementoj de malĉefa strukturo stabiligita de diversaj specoj de interagoj, en kiuj hidrofobaj interagoj ludas gravegan rolon. La stabiligo de la terciara strukturo implikas:
- kovalentaj ligoj (inter la du cisteinaj restaĵoj - pontoj disulfidaj),
- ionikaj ligoj inter kontraŭvole ŝarĝitaj flankaj grupoj de aminoacidaj restaĵoj,
- hidrogenaj ligoj
- hidrofobaj interagoj. Kiam interagas kun la ĉirkaŭaj akvaj molekuloj, la proteina molekulo estas faldita tiel, ke la nepolaj flankaj grupoj de aminoacidoj estas izolitaj de la akva solvo, kaj polusaj hidrofilaj flankaj grupoj aperas sur la surfaco de la molekulo.
Studoj pri la principoj de proteina faldado montris, ke estas oportune distingi alian nivelon inter la nivelo de la sekundara strukturo kaj la atom-spaca strukturo - la faldebla motivo (arkitekturo, struktura motivo). La stila motivo estas determinita per reciproka aranĝo de la sekundaraj strukturoj (α-helicoj kaj β-ŝnuroj) ene de la proteina domajno - kompakta globulo, kiu povas ekzisti memstare aŭ esti parto de pli granda proteino kune kun aliaj domajnoj. Konsideru, ekzemple, unu el la karakterizaj motivoj de la strukturo de proteinoj. La globula proteino montrita en la figuro dekstre, triosofosfatisomerase, havas faldeblan motivon nomatan α / β-cilindro: 8 paralelaj β-ŝnuroj formas β-cilindron ene de alia cilindro kunmetita de 8 α-helicoj. Ĉi tiu motivo troviĝas en ĉirkaŭ 10% de proteinoj.
Estas sciate, ke stilaj motivoj estas sufiĉe konservativaj kaj troviĝas en proteinoj, kiuj havas nek funkciajn, nek evoluciajn rilatojn. Determino de stilaj motivoj baziĝas sur la fizika aŭ racia klasifiko de proteinoj (kiel CATH aŭ SCOP).
Por determini la spacan strukturon de la proteino, metodoj de analizo de difrakto de radiografioj, nuklea magneta resono kaj iuj specoj de mikroskopio estas uzataj.
Kvaternara strukturo
La kvaternara strukturo (aŭ subuneco, domajno) estas la reciproka ordigo de pluraj polipeptidaj ĉenoj en unuopa proteina komplekso.La proteinaj molekuloj, kiuj konsistigas la proteinon kun la kvaternara strukturo, estas formitaj aparte sur la ribosomoj kaj nur post la fino de sintezo ili formas komunan supramolekulan strukturon. Kvaternara proteino povas enhavi ambaŭ identajn kaj malsamajn polipeptidajn ĉenojn. La stabiligo de la Kvaternara strukturo implikas la samajn specojn de interagoj kiel en la stabiligo de la Terciario. Supramolekulaj proteinaj kompleksoj povas konsisti el dekduoj da molekuloj.
Klasifiko laŭ speco de konstruaĵo
Proteinoj povas esti dividitaj en tri grupojn laŭ la ĝenerala speco de strukturo:
- Fibrilaj proteinoj - formas polimerojn, ilia strukturo estas kutime tre regula kaj estas subtenata ĉefe de interagoj inter diversaj ĉenoj. Ili formas mikrofilamentojn, mikrotubulojn, fibrilojn kaj subtenas la strukturon de ĉeloj kaj histoj. Fibrilaj proteinoj inkluzivas keratinon kaj kolagenon.
- Globaj proteinoj estas akvorezistaj, la ĝenerala formo de la molekulo estas pli-malpli sfera.
- Membranproteinoj - havas domojn interplektantajn la ĉelan membranon, sed partoj el ili protrudas de la membrano en la intercelan medion kaj la ĉelan citoplasmon. Membranaj proteinoj agas kiel riceviloj, tio estas, ili transdonas signalojn kaj ankaŭ provizas transmembran transporton de diversaj substancoj. Proteinaj transportiloj estas specifaj, ĉiu el ili pasigas nur certajn molekulojn aŭ certan specon de signalo tra la membrano.
Proteinoj simplaj kaj kompleksaj
Krom peptidaj ĉenoj, multaj proteinoj ankaŭ inkluzivas ne-aminoacidajn grupojn, kaj laŭ ĉi tiu kriterio proteinoj estas dividitaj en du grandajn grupojn - simplajn kaj kompleksajn proteinojn (proteidojn). Simplaj proteinoj konsistas nur el polipeptidaj ĉenoj, kompleksaj proteinoj ankaŭ enhavas ne-aminoacidajn, aŭ prostetajn, grupojn. Depende de la kemia naturo de la prostetaj grupoj, la sekvaj klasoj distingiĝas inter kompleksaj proteinoj:
- Glicoproteinoj enhavantaj kovalentajn karbonhidratajn restaĵojn kiel prostetika grupo, glicoproteinoj enhavantaj mucopolysakaridajn restaĵojn apartenas al la subklaso de proteoglicanoj. En la formado de ligo kun restaĵoj de karbonhidratoj, kutime enhavas hidroksilajn grupojn de serino aŭ treonino. Plej multaj eksterĉelaj proteinoj, precipe imunoglobulinoj, estas glicoproteinoj. En proteoglicanoj, la karbonhidrata parto estas
95% de la tuta maso de la proteina molekulo, ili estas la ĉefa ero de la intercelula matrico,
2. La biologia signifo de la reprodukto de organismoj. Metodoj de reprodukto.
1. Reproduktado kaj ĝia signifo.
Reproduktado - reprodukto de similaj organismoj, kiu havigas
la ekzisto de specioj dum multaj jarmiloj kontribuas al pliiĝo en
la nombro de individuoj de la speco, la kontinueco de la vivo. Sekseca, seksa kaj
vegetativa disvastigo de organismoj.
2. Sekseca reproduktado estas la plej antikva maniero. En
unu organismo okupiĝas pri sekseco, dum plej ofte partoprenas sekso
du individuoj. En plantoj, neseksa reproduktado per sporoj - unu
specialaj ĉeloj. Propagado per sporoj de algoj, musko, ĉevalvosto,
ploregoj, filikoj. Rash de sporoj de plantoj, ilia ĝermado kaj evoluo de
ili novaj filiaj organismoj en favoraj kondiĉoj. Morto de grandega nombro
disputo falanta en malfavorajn kondiĉojn. Malalta probablo de okazo
novaj organismoj el sporoj ĉar ili enhavas malmultajn nutraĵojn kaj
la plantido absorbas ilin ĉefe el la medio.
3. Vegeta disvastigo - disvastigo de plantoj kun
uzante vegetalajn organojn: aeraj aŭ subteraj ŝosoj, partoj de la radiko,
folio, tubero, bulboj. Partopreno en la vegetala disvastigo de unu organismo
aŭ ĝiajn partojn. La afineco de la filina planto kun la patrino, kiel ĝi
daŭrigas la disvolviĝon de la patrina korpo. Bonega efikeco kaj
la disvastiĝo de vegetala disvastiĝo en la naturo, kiel filia organismo
formiĝis pli rapide de la patrina parto ol de la sporo. Vegetaj Ekzemploj
bredado: uzante rizomojn - lilio de la valo, mento, tritiko, ktp., enradikiĝo
malsupraj branĉoj tuŝantaj la grundon (manteloj) - riboj, sovaĝaj vinberoj, lipharoj
- fragoj, bulboj - tulipo, narciso, kroko. Uzo de vegetaĵaro
reproduktiĝante kultivataj plantoj: terpomoj estas propagataj de tuberoj,
bulboj - cepoj kaj ajloj, manteloj - riboj kaj groseroj, radiko
idaro - ĉerizo, pruno, tranĉoj - fruktarboj.
4. Seksa reprodukto. La esenco de seksa reproduktado
en la formado de ĝermaj ĉeloj (gametoj), la fandado de la vira ĝemela ĉelo
(spermo) kaj ino (ovo) - fekundigo kaj disvolviĝo de nova
filina organismo el fekundigita ovo. Dank 'al fekundigo
filia organismo kun pli diversa aro de kromosomoj, kio signifas kun pli
diversaj heredaj trajtoj, rezulte de kiuj ĝi povas esti
pli adaptita al la vivmedio. La ĉeesto de seksa reproduktado en
algoj, muskoj, filikoj, gimnospermoj kaj angiospermoj. Komplikaĵo
la seksa procezo en plantoj dum ilia evoluo, la aspekto de la plej kompleksa
formiĝas en semoplantoj.
5. Disvastigado de semoj okazas helpe de semoj,
ĝi estas karakteriza por gimnospermoj kaj angiospermoj (angiospermoj)
vegetaĵa disvastiĝo ankaŭ estas vasta). Sekvenco de paŝoj
sema disvastigo: polenado - translokigo de poleno al la stigmo de pistilo, ĝia
ĝermado, ekapero dividante du spermojn, ilia progreso en
ovo, poste kunfandiĝo de unu spermo kun ovo, kaj de la alia kun
malĉefa kerno (en angiospermoj). Formado de ovo-semoj -
la embrio kun provizo de nutraĵoj, kaj de la muroj de la ovario - la feto. Semo -
la ĝermo de nova planto, en favoraj kondiĉoj, elkreskas kaj unuafoje
plantido estas nutrata de la nutraĵoj de la semo, kaj tiam ĝiaj radikoj
komencu sorbi akvon kaj mineralojn el la grundo, kaj la folioj - karbona dioksido
gaso el la aero en la sunlumo. La sendependa vivo de nova planto.
Biofiziko de proteinoj
Fizikaj proprietoj de la proteino en la ĉelo, konsiderante la akvan membranon kaj amasiĝon de makromolekuloj (eng.) tre komplika. La hipotezo de proteino kiel ordema "kristala simila sistemo" - "aperioda kristalo" - estas subtenata de analizo de difraktado de radioj X (ĝis rezolucio de 1 angstromo), alta enpakiga denseco, kunlaboro de la denaturacia procezo kaj aliaj faktoj.
En favoro de alia hipotezo, la likvaj similaj ecoj de proteinoj en la procezoj de intraglobulaj movadoj (modelo de limigita saltado aŭ kontinua disvastigo) estas evidentigitaj per eksperimentoj pri disĵetado de neŭtronoj, spektroskopio de Mössbauer.

Universala metodo: ribosoma sintezo
Proteinoj estas sintezitaj de vivaj organismoj el aminoacidoj bazitaj sur informo kodita en genoj. Ĉiu proteino konsistas el unika sinsekvo de aminoacidaj restaĵoj, kiu estas determinita de la nuklea sekvenco de la geno kodanta la proteinon. La genetika kodo estas metodo traduki la nukleotidan sekvencon de DNA (per RNA) en la aminoacidan sekvencon de polipeptida ĉeno. Ĉi tiu kodo determinas la korespondadon de trinucleotidaj sekcioj de RNA, nomataj kodonoj, kaj certajn aminoacidojn, kiuj estas inkluzivitaj en la proteino: la sekvenco de nucleotidoj AUG, ekzemple, respondas al metionino. Ĉar DNA konsistas el kvar specoj de nukleotidoj, la tuta nombro de eblaj kodonoj estas 64, kaj ĉar 20 aminoacidoj estas uzataj en proteinoj, multaj aminoacidoj estas determinitaj de pli ol unu kodono. Tri kodonoj estas sensignivaj: ili servas kiel haltaj signaloj por la sintezo de la polipeptida ĉeno kaj estas nomataj terminaj kodonoj, aŭ haltaj kodonoj.
Genoj kodantaj proteinojn unue estas transskribitaj en la nukleotidan sekvencon de mesaĝaj RNA (mRNA) de RNA-polimeraza enzimoj. En la granda plimulto de kazoj, la proteinoj de vivaj organismoj estas sintezitaj sur ribosomoj - multikomponaj molekulaj maŝinoj prezencaj en la citoplasmo de ĉeloj. La procezo de sintezi polipeptidan ĉenon per ribosomo sur mRNA-matrico nomiĝas tradukado.
Ribosomal proteina sintezo estas esence la sama en prokariotoj kaj eŭkariotoj, sed malsamas en iuj detaloj. Tiel, prokariota mRNA povas esti legata de ribosomoj en la aminoacida sinsekvo de proteinoj tuj post transskribo aŭ eĉ antaŭ ĝia finiĝo. En eŭkariotoj, la primara transskribo unue devas trairi serion de modifoj kaj translokiĝi en la citoplasmon (al la loko de la ribosomo), antaŭ ol traduko povas komenciĝi. La procento de proteina sintezo estas pli alta en procariotoj kaj povas atingi 20 aminoacidojn por dua.

Eĉ antaŭ la komenco de tradukado, aminoacil-tRNA-sintaksaj enzimoj specife ligas aminoacidojn al sia responda transporta RNA (tRNA). Regiono de tRNA, nomata antikodono, povas komplemente pariĝi kun mRNA-kodono, tiel certigante la inkluzivon de aminoacida restaĵo ligita al tRNA en la polipeptida ĉeno konforme al la genetika kodo.
Dum la komenca etapo de tradukado, iniciato, la iniciatilo (kutime metionina) kodono estas rekonita per la malgranda subuneco de la ribosomo, al kiu aminoacilata metionina tRNA estas ligita uzante proteinajn iniciatojn. Post rekoni la komencan kodonon, la granda subuneco kunigas la malgrandan subunecon de la ribosomo, kaj la dua etapo de tradukado, la elongigo, komenciĝas. Ĉe ĉiu paŝo de la ribosomo de la 5'- ĝis 3'-fino de la mRNA, unu kodono estas legata formante hidrogenajn ligojn inter ĝi kaj la transportan RNA-komplementon al ĝi, al kiu estas ligita la responda aminoacida restaĵo. La formado de peptida ligo inter la lasta aminoacida restaĵo de la kreskanta peptido kaj la aminoacida restaĵo ligita al tRNA estas katalizita per ribosoma RNA (rRNA), kiu formas la peptidil-transferasa centro de la ribosomo. Ĉi tiu centro poziciigas la nitrogenajn kaj karbonajn atomojn en pozicio favora por la paŝo de la reago. La tria kaj fina etapo de tradukado, finaĵo, okazas kiam la ribosomo atingas la haltan kodonon, post kio proteinaj finaĵaj faktoroj hidrolizas la ligon inter la lasta tRNA kaj la polipeptida ĉeno, ĉesigante ĝian sintezon. En ribosomoj, proteinoj ĉiam estas sintezitaj de la finaĵo N- ĝis C-finaĵo.
Neribosomal-sintezo
En pli malaltaj fungoj kaj iuj bakterioj, aldona (ne-ribosoma, aŭ multienzima) metodo de biosintezo de peptidoj, kutime de malgranda kaj nekutima strukturo, estas konata.La sintezo de ĉi tiuj peptidoj, kutime malĉefaj metabolitoj, estas efektivigita de proteina komplekso kun alta molekula pezo, NRS-sintasio, sen rekta partopreno de ribosomoj. NRS-sintasezo kutime konsistas el pluraj domajnoj aŭ individuaj proteinoj, kiuj elektas aminoacidojn, formas peptidan ligon kaj liberigas sintezitan peptidon. Ĉi tiuj domajnoj kune formas la modulon. Ĉiu modulo certigas la inkluzivon de unu aminoacido en la sintezita peptido. NRS-sintazoj povas do esti kunmetitaj de unu aŭ pluraj moduloj. Foje, ĉi tiuj kompleksoj inkluzivas domajnon kapablan izomerigi L-aminoacidojn (normalan formon) en D-formon.
Kemia sintezo
Mallongaj proteinoj povas esti sintezitaj kemie per metodoj de organika sintezo, ekzemple kemia ligado. Plej ofte, kemia sintezo de la peptido okazas en la direkto de la C-finaĵo al la N-finaĵo, male al ribosoma biosintezo. La metodo de kemia sintezo produktas mallongajn imunogenajn peptidojn (epitopoj), kiuj tiam estas injektitaj al bestoj por akiri specifajn antikorpojn aŭ hibridomojn. Krome, ĉi tiu metodo estas uzata ankaŭ por akiri inhibilojn de certaj enzimoj. Kemia sintezo permesas la enkondukon de aminoacidaj restaĵoj, kiuj ne troviĝas en konvenciaj proteinoj, ekzemple tiuj kun fluoreskaj etiketoj ligitaj al iliaj flankaj ĉenoj. Kemiaj metodoj por sintezo de proteinoj havas plurajn limojn: ili estas neefikaj kun proteino-longo de pli ol 300 aminoacidaj restaĵoj, artefarite sintezitaj proteinoj povas havi malregulan terciaran strukturon kaj mankas de karakterizaj post-transiraj modifoj.
Post-traduka modifo

Post kiam tradukado estas kompleta, plej multaj proteinoj suferas pliajn kemiajn modifojn nomitajn post-tradukaj modifoj. Pli ol ducent variantoj de post-tradukaj modifoj de proteinoj estas konataj.
Posttransigaj modifoj povas reguligi la vivdaŭron de proteinoj en la ĉelo, ilian enzimatan agadon kaj interagojn kun aliaj proteinoj. En iuj kazoj, post-transiraj modifoj estas deviga etapo de maturiĝo de proteinoj, alie ĝi funkcie senaktiva. Ekzemple, kun la maturiĝo de insulino kaj iuj aliaj hormonoj, necesas limigita proteolizo de la polipeptida ĉeno, kaj kun la maturiĝo de plasmaj membranaj proteinoj, glicosilado estas bezonata.
Posttradukaj modifoj povas esti kaj tre maloftaj kaj maloftaj, ĝis unikaj. Ekzemplo de universala modifo estas ubikvitado (ligado de ĉeno de pluraj molekuloj de la mallonga ubiquitina proteino al proteino), kiu servas kiel signalo por la dispecigo de ĉi tiu proteino fare de la proteasomo. Alia ofta modifo estas glicosilado - oni taksas, ke ĉirkaŭ duono de homaj proteinoj estas glukoksilata. Maloftaj modifoj inkluzivas tirosinigon / detirozinadon kaj poliglikiladon de tubulino.
Unu kaj la sama proteino povas suferi multajn modifojn. Do, histonoj (proteinoj kiuj estas parto de kromatino en eŭkariotoj) sub malsamaj kondiĉoj povas suferi pli ol 150 malsamajn modifojn.
Post-tradukaj modifoj dividiĝas en:
- ĉefaj cirkvitaj modifoj,
- dispecigo de N-fina finaĵa metionina restaĵo,
- limigita proteolizo - forigo de fragmento de proteino, kiu povas okazi de la ekstremoj (disigo de signalaj sekvencoj) aŭ, en iuj kazoj, meze de la molekulo (maturiĝo de insulino),
- la alligiteco de diversaj kemiaj grupoj al liberaj amino kaj karboxilaj grupoj (N-akilado, miristoilado ktp),
- modifoj de flankaj ĉenoj de aminoacidoj,
- la aldono aŭ dispecigo de malgrandaj kemiaj grupoj (glicosilado, fosforiligo, ktp.),
- la aldono de lipidoj kaj hidrokarbonoj,
- ŝanĝo de normaj aminoacidaj restaĵoj al ne-normaj (la formado de citrulino),
- la formado de disulfidaj pontoj inter restaĵoj de cisteino,
- la aldono de malgrandaj proteinoj (sumoylation kaj ubiquitination).
Interĉela transporto kaj ordigo
La proteinoj sintezitaj en la citoplasmo de eŭkariota ĉelo devas esti transportataj al malsamaj ĉelaj organoidoj: la kerno, mitokondrio, endoplasma retikulo (EPR), Golgi-aparato, lisosomoj, ktp., Kaj iuj proteinoj devas eniri la eksterĉelan medion. Por eniri certan sekcion de la ĉelo, la proteino devas havi specifan etikedon. Plejofte tia etikedo estas parto de la aminoacida sinsekvo de la proteino mem (peptido ĉefo, aŭ signala sekvenco de la proteino), sed en iuj kazoj, la oligosakaridoj ligitaj al la proteino estas la etiketo.
La transporto de proteinoj en la EPR efektiviĝas, kiam ili estas sintezitaj, ĉar ribosomoj sintezantaj proteinojn kun signala sekvenco por la EPR "sidas" sur specialaj proteinoj sur ĝia ekstera membrano. De la EPR al la Golgi-aparato, kaj de tie al la lisosomoj kaj al la ekstera membrano aŭ al la eksterĉela mezo, proteinoj eniras vesikan transporton. Proteinoj kun nuklea lokaliza signalo eniras la nukleon tra nukleaj poroj. En mitokondrioj kaj kloroplastoj, proteinoj posedantaj la respondajn signalajn sekvencojn eniras tra specifaj poro-tradukaj poroj kun la partopreno de chaperonoj.
Subteni strukturon kaj degeneron
Subteni ĝustan spacan strukturon de proteinoj estas kerna por ilia normala funkciado. Malĝusta faldiĝo de proteinoj kondukantaj al ilia agregado povas esti kaŭzita de mutacioj, oksido, streskondiĉoj aŭ tutmondaj ŝanĝoj en la fiziologio de la ĉelo. Agregado de proteinoj estas karakteriza signo de maljuniĝo. Oni opinias, ke netaŭga proteina faldado estas la kaŭzo aŭ pligravigado de malsanoj kiel kista fibrozo, lisosoma akumula malsano. same kiel neurodegenerativaj malordoj (Alzheimer, Huntington kaj Parkinson).
En la procezo de ĉela evoluo, kvar ĉefaj mekanismoj estis evoluigitaj por kontraŭstari proteinan agregadon. La unuaj du - ripeta faldado (reverŝado) helpe de chaperonoj kaj dispecigo kun proteazoj - troviĝas ambaŭ en bakterioj kaj en pli altaj organismoj. Aŭtofagio kaj amasiĝo de neĝuste falditaj proteinoj en specialaj ne-membranaj organeloj estas karakterizaj de eŭkariotoj.

La kapablo de proteinoj restarigi la ĝustan tridimensian strukturon post denaturado permesis al ni hipotezi, ke ĉiuj informoj pri la fina strukturo de la proteino estas en sia aminoacida sinsekvo. Nuntempe la teorio ke la stabila konformado de proteino havas minimuman liberan energion estas komparata kun aliaj eblaj konformadoj de ĉi tiu polipeptido.
En ĉeloj ekzistas grupo de proteinoj, kies funkcio estas certigi la ĝustan faldadon de aliaj proteinoj post ilia sintezo sur la ribosomo, restarigi la strukturon de proteinoj post ilia damaĝo, kaj ankaŭ krei kaj disiĝi proteinajn kompleksojn. Ĉi tiuj proteinoj nomiĝas chaaperonoj. La koncentriĝo de multaj ĉafonoj en la ĉelo pliiĝas per akra kresko de la medio-temperaturo, do ili apartenas al la grupo Hsp (angle heat shock proteinoj - varmegaj proteinoj). La graveco de la normala funkciado de ĉperonoj por la funkciado de la korpo povas esti ilustrita per la ekzemplo de α-kristala ĉaperono, kiu estas parto de la lenso de la homa okulo. Mutacioj en ĉi tiu proteino kondukas al nubigado de la lenso pro agregado de proteinoj kaj rezulte al kataraktoj.
Se la tercia strukturo de proteinoj ne povas esti restarigita, ili estas detruitaj de la ĉelo. Enzimoj, kiuj degeneris proteinojn, estas nomataj proteazoj.En la loko de atako de la substrata molekulo, proteolitikaj enzimoj dividiĝas al endopeptidasoj kaj exopeptidasoj:
- Endopeptidasoj, aŭ proteinase, fendetiĝas peptidajn ligojn ene de la peptida ĉeno. Ili rekonas kaj ligas mallongajn peptidajn sinsekvojn de substratoj kaj relative specife hidrolizas la ligojn inter certaj aminoacidaj restaĵoj.
- Exopeptidases hidrolizas peptidojn de la ekstremoj de la ĉeno: aminopeptidasoj de la N-finaĵo, karboxokseptidasoj de la C-termino. Fine dipeptidasoj disiĝas nur dipeptidoj.
Laŭ la kataliza mekanismo, la Internacia Unio por Biokemio kaj Molekula Biologio identigas plurajn klasojn de proteazoj, inkluzive de serinaj proteazoj, aspartaj proteazoj, cisteine-proteazoj kaj metaloproteasoj.
Speciala tipo de proteazoj estas la proteasomo, granda multisubunita protezo ĉeestanta en la kerno kaj citoplasmo de eŭkariotoj, arkaea, kaj iuj bakterioj.
Por ke la proteazoma celo estu fendita de la proteino, ĝi devas esti etikedita ligante al ĝi malgrandan ubiquitin-proteinon. La aldona reago de ubiquitino estas katalizita de la enzimoj ubiquitin ligases. La aldono de la unua ubiquitina molekulo al la proteino servas kiel signalo por ligasoj por plua aldono de ubiquitinaj molekuloj. Rezulte, polubikitina ĉeno estas ligita al la proteino, kiu ligiĝas al la proteasomo kaj certigas fendadon de la cela proteino. Ĝenerale, ĉi tiu sistemo nomiĝas ubiquitin-dependa proteina degenero. La degradado de 80-90% de intracelaj proteinoj okazas kun la partopreno de la proteasomo.
Proteina degenero en perksisomoj gravas por multaj ĉelaj procezoj, inkluzive de la ĉela ciklo, regulado de gena esprimo, kaj respondo al oksida streso.

Aŭtofagio estas la procezo de degradado de longvivaj biomolekuloj, precipe proteinoj, same kiel organeloj en lisosomoj (ĉe mamuloj) aŭ vakuoloj (en feĉo). Aŭtofagio akompanas la esencan agadon de iu normala ĉelo, sed la manko de nutraĵoj, la ĉeesto de difektitaj organeloj en la citoplasmo kaj, fine, la ĉeesto de parte denaturigitaj proteinoj kaj iliaj agregoj en la citoplasmo povas servi kiel stimuloj por plibonigi la procezojn de aŭtofagio en ĉeloj.
Tri specoj de aŭtofagio distingiĝas: mikroautofagia, makrohaŭtofagio, kaj ĉero-dependa aŭtofagio.
Dum mikroautofagio, makromolekuloj kaj fragmentoj de ĉelaj membranoj estas kaptitaj de la lisosomo. Tiel la ĉelo povas digesti proteinojn kun manko de energio aŭ konstrumaterialo (ekzemple dum malsato). Sed la procezoj de mikroautofagio okazas sub normalaj kondiĉoj kaj ĝenerale estas nediskriminataj. Foje ankaŭ organoidoj estas digestitaj dum mikroautofagio, ekzemple mikroautofagio de perksisomoj kaj parta mikroautofagio de kernoj, en kiuj la ĉelo restas farebla, estas priskribita en feĉo.
En makroautofagio, parto de la citoplasmo (ofte enhavas iujn ajn organoidojn) estas ĉirkaŭita de membrana kupeo simila al cisterno de la endoplasma retikulo. Rezulte tiu retejo estas apartigita de la resto de la citoplasmo per du membranoj. Tiaj du-membranaj organeloj nomiĝas aŭtofagosomoj. Aŭtofagozomoj kunfandiĝas kun lisosomoj, formante aŭtofagolizosomojn, en kiuj estas digestitaj la organeloj kaj la ceteraj enhavoj de aŭtofagojomoj. Ŝajne makrohaŭtofagio ankaŭ estas ne selektema, kvankam oni ofte emfazas, ke helpe de ĝi la ĉelo povas liberiĝi de organoidoj "malaktuaj" (mitokondrioj, ribosomoj, ktp).
La tria speco de aŭtofagio dependas de chaperono. En ĉi tiu metodo, direktita transporto de parte denaturigitaj proteinoj el la citoplasmo tra la lisosoma membrano al ĝia kavo okazas, kie ili estas digestitaj. Ĉi tiu tipo de aŭtofagio, priskribita nur ĉe mamuloj, estas induktita de streso.

JUNQ kaj IPOD
Sub streso, kiam eŭkariota ĉelo ne povas fari fronte al la amasiĝo de multnombraj proteinoj denaturitaj, ili povas esti senditaj al unu el du specoj de provizoraj organeloj - JUNQ kaj IPOD (angla) ruso. .
JUNQ (angle: JUxta Nuclear Quality control control compartiment) estas asociita kun la ekstera flanko de la nuklea membrano kaj enhavas ubiquitinitajn proteinojn, kiuj povas rapide translokiĝi al la citoplasmo, same kiel chaperonoj kaj proteasomoj. La celita funkcio de JUNQ estas refolde kaj / aŭ degradi proteinojn.
IPOD (angle Insoluble Protein Deponejo - loko de deponejo de nesolveblaj proteinoj) situas proksime de la centra vakuolo kaj enhavas senmovajn agregojn de amiloid-formantaj proteinoj. La amasiĝo de ĉi tiuj proteinoj en la IPOD povas malhelpi ilian interagadon kun normalaj ĉelaj strukturoj, tial oni kredas, ke ĉi tiu inkludo havas protektan funkcion.
La funkcioj de proteinoj en la korpo
Kiel aliaj biologiaj makromolekuloj (polisakaridoj, lipidoj kaj nukleaj acidoj), proteinoj estas esencaj komponantoj de ĉiuj vivaj organismoj kaj ludas gravan rolon en la vivo de la ĉelo. Proteinoj efektivigas metabolajn procezojn. Ili estas parto de intracelulaj strukturoj - organeloj kaj citoskeleto, sekreciitaj en la eksterĉelan spacon, kie ili povas agi kiel signalo transdonita inter ĉeloj, partopreni la hidrolizon de manĝaĵoj kaj la formado de intercelula substanco.
La klasifiko de proteinoj laŭ iliaj funkcioj estas iom arbitra, ĉar la sama proteino povas plenumi plurajn funkciojn. Bone studita ekzemplo de tia multfunkcieco estas lisila tRNA-sintezo, enzimo el la klaso de aminoacilaj tRNA-sintazoj, kiu ne nur ligas la lisinan restaĵon al tRNA, sed ankaŭ reguligas la transskribon de pluraj genoj. Proteinoj plenumas multajn funkciojn pro sia enzimata aktiveco. Do, la enzimoj estas miozina motora proteino, reguliga proteino kinasa proteino, transportas proteinon natrio-kalio-adenosina trifosfatase, ktp.

Kataliza funkcio
La plej konata funkcio de proteinoj en la korpo estas la katalizo de diversaj kemiaj reagoj. Enzimoj estas proteinoj, kiuj havas specifajn katalizajn proprietojn, tio estas, ke ĉiu enzimo katalizas unu aŭ plurajn similajn reagojn. Enzimoj katalizas la disigon de kompleksaj molekuloj (katabolismo) kaj ilian sintezon (anabolismo), inkluzive de replikado kaj riparo de DNA kaj sintezo de RNA-matrico. Antaŭ 2013, pli ol 5.000 enzimoj estis priskribitaj. La akcelo de la reago rezulte de enzimata katalizo povas esti enorma: la reago katalizita de la enzimo orotidin-5'-fosfata decarboxilasa, ekzemple, procedas 10 17 fojojn pli rapide ol la ne katalizita (la duonvivo de la dekarboxilado de orotika acido estas 78 milionoj da jaroj sen la enzimo kaj 18 milisekundoj). Molekuloj, kiuj kuniĝas al la enzimo kaj ŝanĝiĝas rezulte de la reago, estas nomataj substratoj.
Kvankam enzimoj kutime konsistas el centoj da aminoacidaj restaĵoj, nur malgranda parto el ili interagas kun la substrato, kaj eĉ pli malgrandaj kvantoj - averaĝe 3-4 aminoacidaj restaĵoj, ofte lokitaj for unu de la alia en la primara strukturo - estas rekte implikitaj en katalizo. La parto de la enzima molekulo kiu provizas la ligadon de la substrato kaj katalizo estas nomata aktiva centro.
En 1992, la Internacia Unio pri Biokemio kaj Molekula Biologio proponis la finan version de la hierarkia nomenklaturo de enzimoj laŭ la speco de reagoj katalizitaj de ili. Laŭ ĉi tiu nomaro, la nomoj de enzimoj ĉiam devas havi finaĵon -la bazoj kaj formi el la nomoj de la katalizitaj reagoj kaj iliaj substratoj. Al ĉiu enzimo oni atribuas individuan kodon per kiu estas facile determini sian pozicion en la hierarkio de enzimoj.Laŭ la tipo de katalizitaj reagoj, ĉiuj enzimoj estas dividitaj en 6 klasojn:
- CF 1: oksidoreduktaseoj kiuj katalizas redox-reagojn,
- CF 2: Transferaĵoj, kiuj katalizas la translokadon de kemiaj grupoj de unu substrata molekulo al alia,
- CF 3: Hidrolasoj katalizantaj la hidrolizon de kemiaj ligoj,
- CF 4: Laseoj katalizantaj la rompadon de kemiaj ligoj sen hidrolizo kun la formado de duobla ligo en unu el la produktoj,
- CF 5: Izomerasoj, kiuj katalizas strukturajn aŭ geometriajn ŝanĝojn en la substrata molekulo,
- CF 6: Ligazoj, kiuj katalizas la formadon de kemiaj ligoj inter substratoj pro hidrolizo de la liga ATP-difosfato aŭ simila trifosfato.
Struktura funkcio
La strukturaj proteinoj de la citoskeleto, kiel speco de armaturaĵo, donas formon al ĉeloj kaj multaj organoidoj kaj okupiĝas pri ŝanĝo de la formo de ĉeloj. Plej multaj strukturaj proteinoj estas filamentaj: aktino kaj tubulaj monomeroj estas, ekzemple, globulaj, solveblaj proteinoj, sed post polimerigo ili formas longajn ŝnurojn, kiuj konsistigas la citoskeleton, kio permesas al la ĉelo konservi formon. Kolageno kaj elastino estas la ĉefaj komponentoj de la intercelula substanco de konektiva histo (ekzemple kartilago), kaj haroj, ungoj, birdaj plumoj kaj iuj ŝeloj estas formitaj de alia keratina struktura proteino.

Protekta funkcio
Estas pluraj specoj de protektaj funkcioj de proteinoj:
- Fizika protekto. Fizika protekto de la korpo estas provizita de kolageno, proteino, kiu estas la bazo de la intercelula substanco de konekteblaj histoj (inkluzive de ostoj, kartilago, tendenoj kaj profundaj tavoloj de la haŭto (dermo)), keratino, kiu formas la bazon de kornaj ŝildoj, haroj, plumoj, kornoj kaj aliaj derivaĵoj de la epidermo. Tipe, tiaj proteinoj estas konsiderataj kiel proteinoj kun struktura funkcio. Ekzemploj de proteinoj de ĉi tiu grupo estas fibrinogeno kaj trombino implikita en sanga koaguliĝo.
- Kemia protekto. La ligado de toksinoj al proteinaj molekuloj povas havigi ilian sentoksiĝon. Precipe grava rolo en la sentoksiĝo ĉe homoj ludas hepatajn enzimojn, kiuj detruas venenojn aŭ konvertas ilin en solveblan formon, kio kontribuas al ilia rapida forigo el la korpo.
- Imuna defendo. Proteinoj, kiuj konsistigas sangon kaj aliajn korpajn fluidojn, estas implikitaj en la defenda respondo de la korpo al ambaŭ damaĝo kaj atako de patogenoj. Proteinoj de la komplementa sistemo kaj antikorpoj (imunoglobulinoj) apartenas al la proteinoj de la dua grupo, ili neŭtraligas bakteriojn, virusojn aŭ fremdajn proteinojn. Antikorpoj, kiuj estas parto de la adapta imuna sistemo, ligiĝas al substancoj fremdaj al la korpo, antigenoj, kaj tiel neŭtraligas ilin, direktante ilin al la lokoj de detruo. Antikorpoj povas esti sekreciaj en la intercelan spacon aŭ fiksiĝi en la membranoj de specialigitaj B-limfocitoj nomataj plasmocitoj.
Reguliga funkcio
Multaj procezoj en la ĉeloj estas reguligitaj per proteinaj molekuloj, kiuj servas nek kiel fonto de energio nek kiel konstruaj materialoj por la ĉelo. Ĉi tiuj proteinoj reguligas ĉelan antaŭeniĝon en la ĉela ciklo, transskribo, tradukado, dismetado, la agado de aliaj proteinoj, kaj multaj aliaj procezoj. Proteinoj plenumas la reguligan funkcion aŭ pro enzimata agado (ekzemple proteinoj kinaseoj), aŭ pro specifa ligado al aliaj molekuloj. Tiel, transkripcifaktoroj, aktivigaj proteinoj kaj subpremaj proteinoj, povas reguligi la transskriban intenson de genoj ligante sin al iliaj reguligaj sekvencoj. Je traduka nivelo, la legado de multaj ARNm estas ankaŭ reguligita per la aldono de proteinaj faktoroj.
La plej grava rolo en la regulado de intracelulaj procezoj ludas proteinajn kinaseojn kaj proteinajn fosfatazojn - enzimojn, kiuj aktivigas aŭ malhelpas la agadon de aliaj proteinoj ligante ilin aŭ dividante fosfatajn grupojn.

Signala funkcio
La funkcia signalo de proteinoj estas la kapablo de proteinoj funkcii kiel signalaj substancoj, transdoni signalojn inter ĉeloj, histoj, organoj kaj organismoj. Ofte la signala funkcio kombiniĝas kun la reguliga, ĉar multaj intracelulaj reguligaj proteinoj ankaŭ transdonas signalojn.
La signa funkcio plenumas hormonajn proteinojn, citokinojn, kreskajn faktorojn ktp.
Hormonoj estas portataj de sango. Plej multaj bestaj hormonoj estas proteinoj aŭ peptidoj. La ligado de la hormono al ĝia ricevilo estas signalo, kiu ekigas ĉelan respondon. Hormonoj reguligas la koncentriĝon de substancoj en la sango kaj ĉeloj, kreskon, reprodukton kaj aliajn procezojn. Ekzemplo de tiaj proteinoj estas insulino, kiu reguligas la koncentriĝon de glukozo en la sango.
Ĉeloj interagas inter si uzante signalajn proteinojn transdonitajn tra la intercelula substanco. Tiaj proteinoj inkluzivas, ekzemple, citokinojn kaj kreskfaktorojn.
Citokinoj estas peptidaj signalaj molekuloj. Ili reguligas interagojn inter ĉeloj, determinas ilian postvivadon, stimulas aŭ malhelpas kreskon, diferencadon, funkcian agadon kaj apoptozon, certigas la kunordigon de la imuna, endokrina kaj nerva sistemo. Ekzemplo de citokinoj estas la faktoro de tumora necrozo, kiu transdonas inflamajn signalojn inter la ĉeloj de la korpo.
Senpaga (standby) funkcio
Tiaj proteinoj inkluzivas la nomitajn rezervajn proteinojn, kiuj estas konservataj kiel fonto de energio kaj substanco en la semoj de plantoj (ekzemple 7S kaj 11S globulinoj) kaj ovoj de bestoj. Multaj aliaj proteinoj estas uzataj en la korpo kiel fonto de aminoacidoj, kiuj siavice estas pioniroj de biologie aktivaj substancoj, kiuj reguligas metabolajn procezojn.

Funkcia ricevilo
Riceviloj de proteinoj troveblas en la citoplasmo kaj integriĝas en la ĉela membrano. Unu parto de la ricevilo-molekulo ricevas signalon, kiu plej ofte servas per kemia substanco, kaj en iuj kazoj - malpeza, mekanika streĉado (ekzemple streĉanta) kaj aliaj stimuloj. Kiam signalo estas elmontrita al specifa parto de la molekulo - la receptoro-proteino - ĝiaj konformaj ŝanĝoj okazas. Rezulte ŝanĝiĝas la konformiĝo de alia parto de la molekulo, kiu transdonas signalon al aliaj ĉelaj komponentoj. Estas pluraj mekanismoj pri transdono de signaloj. Iuj riceviloj katalizas ian kemian reagon, aliaj servas kiel jonoj-kanaloj, kiuj malfermiĝas aŭ fermiĝas per la ago de signalo, dum aliaj specife kunigas molekulojn mediantajn. Ĉe membranaj riceviloj, la parto de la molekulo, kiu ligas al la signal-molekulo, estas sur la surfaco de la ĉelo, kaj la domajno, kiu elsendas la signalon, estas interne.
Motoro (motoro) funkcio
Tuta klaso de motorproteinoj provizas korpajn movadojn, ekzemple muskola kuntiriĝo, inkluzive lokomotivon (miozino), movadon de ĉeloj ene de la korpo (ekzemple amoeboida movado de leŭkocitoj), movadon de cilia kaj flagelo, same kiel aktivan kaj direktitan intracelan transporton (kinesino, dyneino) . Dyneinoj kaj kineinoj transportas molekulojn laŭ mikrotubuloj uzante ATP-hidrolizon kiel energifonton. Dyneinoj translokigas molekulojn kaj organelojn de la ekstercentraj partoj de la ĉelo direkte al centrosomo, kineinoj - en la kontraŭa direkto. Dyneinoj ankaŭ respondecas pri la movado de cilioj kaj flageloj de eŭkariotoj. Citoplasmaj variantoj de miozino povas esti implikitaj en transportado de molekuloj kaj organoidoj tra mikrofilamentoj.
Proteinoj en la metabolo
Plej multaj mikroorganismoj kaj plantoj povas sintezi 20 normajn aminoacidojn, kaj kromajn (ne normajn) aminoacidojn, kiel ekzemple citrulino.Sed se aminoacidoj estas en la medio, eĉ mikroorganismoj konservas energion transportante aminoacidojn al la ĉeloj kaj malŝaltante iliajn biosintetajn vojojn.
Aminoacidoj, kiuj ne povas esti sintezitaj de bestoj, estas nomataj esencaj. Ĉefa enzimo en biosintezaj vojoj, ekzemple, aspartato kinase, kiu katalizas la unuan paŝon en la formado de lizino, metionino kaj treonino el aspartato, forestas ĉe bestoj.
Bestoj ĉefe ricevas aminoacidojn el la proteinoj trovitaj en manĝaĵoj. Proteinoj estas detruitaj dum digesto, kiu kutime komenciĝas per la denaturado de la proteino metante ĝin en acidan medion kaj hidrolizi ĝin uzante enzimojn nomitajn proteazoj. Iuj aminoacidoj akiritaj rezulte de digesto estas uzataj por sintezi korpajn proteinojn, dum la ceteraj konvertiĝas al glukozo dum glukogenozo aŭ uzas en la ciklo de Krebs. La uzo de proteino kiel energifonto gravas precipe en kondiĉoj de fastado, kiam la propraj proteinoj de la korpo, precipe muskoloj, servas kiel energifonto. Aminoacidoj ankaŭ estas grava fonto de nitrogeno en la korpa nutrado.
Ne ekzistas unuigitaj normoj por konsumado de homaj proteinoj. La mikroflora floro de la granda intesto sintezas aminoacidojn, kiujn oni ne konsideras en la preparado de proteinaj normoj.
Studadmetodoj
La strukturo kaj funkcioj de proteinoj estas studataj ambaŭ pri purigitaj preparoj en vitro, kaj en sia natura medio en vivanta organismo, in vivo. Studoj pri puraj proteinoj sub kontrolitaj kondiĉoj estas utilaj por determini iliajn funkciojn: kinetikaj ecoj de la kataliza agado de enzimoj, relativa afineco por diversaj substratoj, ktp. Proteinaj studoj in vivo en ĉeloj aŭ en tutaj organismoj disponigas pliajn informojn pri kie ili funkcias kaj kiel ilia aktiveco estas reguligita.
Molekula kaj ĉela biologio

Molekulaj kaj ĉelaj biologiaj metodoj estas ofte uzataj por studi la sintezon kaj lokalizadon de proteinoj en ĉelo. Metodo por studi lokalizon estas vaste uzata, surbaze de la sintezo de aimera proteino en ĉelo, konsistanta el la studita proteino, konektita al "raportisto", ekzemple, verda fluoreska proteino (GFP). La loko de tia proteino en la ĉelo videblas per fluoreska mikroskopo. Krome, proteinoj videblas uzante antikorpojn, kiuj rekonas ilin, kiuj siavice portas fluoreskan etikedon. Ofte, samtempe kun la studita proteino, ofte konataj proteinoj de tiaj organeloj kiel la endoplasma retikulo, Golgi-aparato, lisosomoj kaj vakuoloj estas videblaj, kio ebligas pli precizan determinon de la lokalizo de la studita proteino.
Immunohistokemiaj metodoj kutime uzas antikorpojn konjugitajn al enzimoj, kiuj katalizas la formadon de lumineska aŭ kolora produkto, kio ebligas vin kompari lokon kaj kvanton de proteino studita en la specimenoj. Pli malofta tekniko por determini la lokon de proteinoj estas la ekvilibra ultracentrifugado de ĉelaj frakcioj en la gradiento de sukerozo aŭ cesio-klorido.
Fine, unu el la klasikaj metodoj estas imunoelektronika mikroskopo, kiu estas esence simila al imunofluoreskenta mikroskopio kun la diferenco, ke oni uzas elektronan mikroskopon. La specimeno estas preparita por elektronika mikroskopio, kaj poste prilaborita per antikorpoj al proteino, kiu estas konektita al elektron-densa materialo, kutime oro.
Uzante retej-mutagenezon, esploristoj povas ŝanĝi la aminoacidan sekvencon de proteino kaj sekve ĝian spacan strukturon, lokon en la ĉelo kaj la reguladon de ĝia agado. Uzante ĉi tiun metodon, uzante modifitajn tRNAojn, artefaritaj aminoacidoj ankaŭ povas esti enkondukitaj en proteino kaj proteinoj kun novaj proprietoj povas esti konstruitaj.
Biokemiaj
Por realigi analizon en vitro la proteino devas esti elpurigita el aliaj ĉelaj komponentoj. Ĉi tiu procezo kutime komenciĝas per la detruo de ĉeloj kaj akirado de la nomata ĉela eltiraĵo. Plue, per centrifugaj kaj ultracentrifugaj metodoj, ĉi tiu eltiraĵo povas esti dividita en: frakcio enhavanta solveblajn proteinojn, frakcion enhavantan membranajn lipidojn kaj proteinojn, kaj frakcion enhavantan ĉelajn organelojn kaj nukleajn acidojn.

Proteina precipitaĵo per salado estas uzata por disigi proteinajn miksaĵojn, kaj ankaŭ permesas proteinan koncentriĝon. Analizo de sedimentado (centrifugado) permesas frakcii proteinajn miksaĵojn per la valoro de la konstanta sedimentado de unuopaj proteinoj, mezurita en svedbergoj (S). Diversaj specoj de kromatografio tiam estas uzataj por izoli la deziratan proteinon aŭ proteinojn bazitajn sur propraĵoj kiel molekula pezo, ŝarĝo kaj afineco. Krome, proteinoj povas esti izolitaj laŭ ilia ŝarĝo uzante elektronofokon.
Por simpligi la procezon de purigo de proteinoj, ofte uzas genetikan inĝenieradon, kio permesas krei derivaĵojn de proteinoj konvenaj por purigo sen tuŝi ilian strukturon aŭ aktivecon. "Etikedoj", kiuj estas malgrandaj aminoacidaj sekvencoj, ekzemple, ĉeno de 6 aŭ pli da histidinaj restaĵoj, kaj estas ligitaj al unu fino de la proteino. Kiam la eltiraĵo de la ĉeloj sintezantaj la "etikeditan" proteinon estas pasita tra kromatografia kolumno enhavanta nikelajn jonojn, histidino ligas kun nikelo kaj restas sur la kolumno, dum la ceteraj komponentoj de la lisato pasas tra la kolumno sen obstakloj (nikel-ĉelata kromatografio). Multaj aliaj etikedoj estis dizajnitaj por helpi esploristojn izoli specifajn proteinojn de kompleksaj miksaĵoj, plej ofte uzante afinec-kromatografion.
La grado de puriga proteino povas esti determinita se ĝia molekula pezo kaj izoelektra punkto estas konataj - uzante diversajn specojn de ĝel-electroforesis - aŭ mezurante la enzimatan agadon se la proteino estas enzimo. Masa spektrometrio permesas identigi la elektitan proteinon per ĝia molekula pezo kaj la maso de ĝiaj fragmentoj.
Proteomikoj
La totalo de ĉelaj proteinoj nomiĝas proteomo, ĝia studo - proteomiko, nomata per analogio kun genomiko. Ŝlosilaj eksperimentaj proteomikoj-metodoj inkluzivas:
- 2D-elektroforesis, kiu permesas la apartigon de multikomponentaj proteinaj miksaĵoj,
- masa spektrometrio, kiu permesas la identigon de proteinoj per la maso de iliaj konstantaj peptidoj kun alta fluo,
- proteinoj microarrays, kiuj permesas samtempe mezuri la enhavon de granda nombro da proteinoj en la ĉelo,
- du-hibrida feĉo-sistemo , kiu permesas vin studi sisteme interrilatojn proteino-proteino.
La totalo de ĉiuj biologie signifaj interagoj de proteinoj en ĉelo estas nomata interaktomo. Sistema studo de la strukturo de proteinoj reprezentantaj ĉiujn eblajn specojn de terciaraj strukturoj estas nomata struktura genomiko.
Strukturo-Antaŭdiro kaj Modeligado
Antaŭdiro de spaca strukturo uzanta komputilajn programojn (en silico) permesas konstrui proteinajn modelojn kies strukturo ankoraŭ ne estis determinita eksperimente. La plej sukcesa speco de struktura prognozo, nomata homologa modelado, dependas de la ekzistanta "ŝablona" strukturo, simila en aminoacida sinsekvo al la modeligita proteino. La metodoj por antaŭdiri la spacan strukturon de proteinoj estas uzataj en la disvolva kampo de genetika inĝenierado de proteinoj, kun la helpo de kiuj jam akiris terciajn strukturojn de proteinoj. Pli kompleksa komputila tasko estas la prognozo de intermolekulaj interagoj, kiel molekula doko kaj prognozo de proteinoj-proteinoj.
La faldeblaj kaj intermolekulaj interagoj de proteinoj povas esti modeligitaj per molekula mekaniko. , precipe, molekula dinamiko kaj la metodo Monte Carlo, kiuj pli kaj pli utiligas paralelan kaj distribuitan komputadon (ekzemple la Folding @ home-projekton).La faldiĝo de malgrandaj α-helicaj proteinaj domajnoj, kiel ekzemple vilina proteino aŭ unu el la HIV-proteinoj, estis sukcese modeligita en silico. Uzante hibridajn metodojn, kiuj kombinas norman molekulan dinamikon kun kvantuma mekaniko, la elektronikaj statoj de la vida pigmenta rodopsino estis esploritaj.

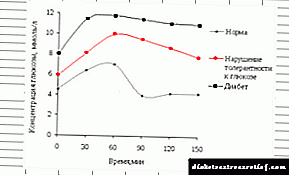
-
Kiel uzi la drogon Biosulin?
Liberiga formo kaj konsisto La drogo havas tri dozajn formojn: Injekta solvo Biosulino R, Suspendo por administrado de s / c Biosulino N, Suspendo por administrado de s / c Biosulin 30/70. ... -

-

-