Reguligo de kolesterola sintezo
La regulado de la sintezo de kolesterolo - ĝia kerna enzimo (HMG-CoA reductase) efektivigas diversajn manierojn.
Fosforilación / defosforilación de HMG-reduktasa. Kun pliigo en la rilatumo insulino / glucagono, ĉi tiu enzimo fosforilas kaj pasas al aktiva stato. La ago de insulino efektiviĝas per 2 enzimoj.
HMG-CoA-reduktasa kinasa fosfatasa, kiu igas la kinaseon neaktiva defosforilata stato:
Fosfotasa HMG-CoA-reduktasa transformante ĝin en defosforilatan aktivan staton. La rezulto de ĉi tiuj reagoj estas la formado de aktiva formo defosforilata de HMG-CoA-reduktasa.
Sekve, dum la absorba periodo, kolesterolo plialtiĝas. Dum ĉi tiu periodo, la havebleco de la komenca substrato por la sintezo de kolesterolo - acetilo - CoA pliiĝas (rezulte de manĝado de manĝaĵoj enhavantaj karbonhidratojn kaj grasojn, ĉar CoA-acetilo formiĝas dum rompo de glukozo kaj grasaj acidoj).
En la postabsorba stato, gluĉagono tra proteingenase A stimulas la fosforiligon de HMG - CoA - reduktase, igante ĝin neaktiva stato. Ĉi tiu ago estas plibonigita per la fakto, ke samtempe glukozono stimulas la fosforiladon kaj senaktivigon de la fosfataza HMG-CoA-reduktase, konservante tiel HMG-CoA-reduktase fosforilatan neaktivan staton. Rezulte de tio, la sintezo de kolesterolo en la postabsorbado kaj dum fastado estas malhelpita de endogena sintezo. Se la kolesterola enhavo en manĝaĵo estis alportita al 2%, tiam la sintezo de endogena kolesterolo malpliiĝis akre. Sed la kompleta ĉeso de sintezo de kolesterolo ne okazas.
La grado de inhibicio de la biosintezo de kolesterolo sub la influo de kolesterolo venanta de manĝaĵo varias de homo al persono. Ĉi tio indikas la individuecon de la procezoj de formiĝo de kolesterolo. Reduktante la intensecon de kolesterola sintezo, eblas redukti ĝian koncentriĝon en sango.
Se la ekvilibro inter la konsumado de kolesterolo kun manĝaĵo kaj ĝia sintezo en la korpo unuflanke kaj la ekskrecio de galaj acidoj kaj kolesterolo aliflanke, la koncentriĝo de kolesterolo en la histoj kaj sango ŝanĝiĝas. La plej gravaj konsekvencoj estas asociitaj kun pliigo de sanga kolesterola koncentriĝo (hipercolesterolemia), dum la verŝajneco disvolvi aterosklerozon kaj kolelitiasis.
Familia hipercolesterolemio (HCS) - ĉi tiu formo estas multe pli ofta - ĉirkaŭ 1 paciento po 200 homoj. Heredita difekto en HCS estas malobservo de la absorbo de LDL de ĉeloj, kaj tial malkresko de la indico de LDL-katabolismo. Rezulte la koncentriĝo de LDL en la sango pliiĝas, same kiel kolesterolo, ĉar estas multe da ĝi en LDL. Tial kun HCS estas karakteriza deponejo de kolesterolo en histoj, precipe en haŭto (xanthomoj), en muroj de arterioj.
Malhelpo de sintezo de HMG-CoA-reduktase
La fina produkto de la metabola vojo de kolesterolo. Ĝi reduktas la ritmon de transskribo de la geno HMG-CoA reduktasa, tiel inhibante sian propran sintezon. La hepato aktive sintezas bilinajn acidojn el kolesterolo, kaj tial la galaj acidoj malhelpas la agadon de la geno HMG-CoA reduktasa. Ĉar HMG-CoA reductase ekzistas post sintezo de ĉirkaŭ 3, inhibicio de la sintezo de ĉi tiu kolesterola enzimo estas efika regulado.
Ĉu vi ne trovis tion, kion vi serĉis? Uzu la serĉon:
Interŝanĝo de esteroj de kolesterolo
La kolesterolo enhavas senpagajn esterojn de kolesterolo kaj kolesterolo, kiuj troviĝas ambaŭ en ĉeloj kaj en sangaj lipoproteinoj.
2a parto. Metabolo kaj energio
En ĉeloj, la esterigo de kolesterolo okazas per la ago de acil-CoA-kolesterolo-aciltransferasa (AChAT):
Acyl-CoA + Kolesterolo - * HS-KoA + Acylcholesterol
En homaj ĉeloj, linoleilkolesterolo estas ĉefe formata. Male al senpaga kolesterolo, ĝiaj esteroj en ĉelmembranoj estas enhavitaj en tre malgrandaj kvantoj kaj troviĝas ĉefe en la citosol kiel parto de lipidaj gutoj. La formado de esteroj povas esti konsiderata, unuflanke, kiel mekanismo por forigi ekscesan kolesterolon de la membranoj, kaj aliflanke, kiel mekanismo por la stokado de kolesterolo en la ĉelo. Rimedo-mobilizo okazas kun partopreno de esterazaj enzimoj, kiuj hidrolizas esterojn de kolesterolo:
Acilkolesterolo + H 2O - * Grasa acido + Kolesterolo
La sintezo kaj hidrolizo de esteroj okazas en multaj ĉeloj, sed estas precipe aktiva en ĉeloj de la suprena cortekso: en ĉi tiuj ĉeloj ĝis 80% de ĉiuj kolesteroloj estas reprezentitaj de esteroj, dum en aliaj ĉeloj kutime malpli ol 20%.
La formado de esteroj en sangaj liponroteinoj okazas kun la partopreno de lecitin-kolesterola aciltransferase (LHAT), kiu katalizas la translokigon de la acila restaĵo el la i-pozicio de lecitino al kolesterolo (Fig. 10.31). LHAT estas formita en la hepato, sekreciita en la sangon kaj ligita al lipoproteinoj. La esteriga indico por malsamaj lipoproteinoj estas signife malsama kaj dependas de la ĉeesto de apolipoproteinoj, kiuj aktivigas LHAT (ĉefe apo-AT, same kiel CI) aŭ inhibicias (C-II) ĉi enzimon.La plej aktiva LHAT en HDL, en kiu apo-AT devas olee 2/3 de ĉiuj proteinoj. La plej granda coli honoroj formis esteroj de oleico kaj linoleico acidoj. La alia lipoproteínas ester formado okazas malpli rapide ol en HDL.
Fig. 10.31. La formado de esteroj de kolesterolo sub la agado de LHAT
LHAT estas lokalizita en la surfaca tavolo de HDL kaj uzas kolesterolon en la fosfolipida monostrato kiel substrato. La kolesteroloj formitaj ĉi tie, pro sia kompleta hidrofobeco, estas nebone konservitaj
Ĉapitro 10. Metabolismo kaj Lipida Funkcio
fosfolipida monokapa kaj enmeta en la lipidan kernon de la lipoproteino. Samtempe oni liberigas lokon por kolesterolo en la fosfolipida monostrato, kiu povas esti plenigita per kolesterolo el ĉelaj membranoj aŭ el aliaj lipoproteinoj. Tiel, HDL ŝajnas esti kolesterolo kaptita de la agado de LHAT.
Bile Acida Sintezo
En la hepato, parto de la kolesterolo estas transformita al galaj acidoj. Gallaj acidoj povas esti konsiderataj kiel derivaĵoj de klanika acido (Fig. 10.32).
Kolanika acido kiel tia ne estas formita en la korpo. En hepatocitoj, kolesterolo rekte produktas chenodeoksikolajn kaj ĉoliajn acidojn - primarajn bilidajn acidojn (Fig. 10.33, vidu ankaŭ Fig. 10.12).
Biosintezo de kolesterolo
Biosintezo de kolesterolo okazas en la endoplasma retikulo. La fonto de ĉiuj karbonaj atomoj en la molekulo estas acetil-SCoA, kiu venas ĉi tie el mitokondrioj kiel parto de citrato, kiel en la sintezo de grasaj acidoj. La kolesterola biosintezo konsumas 18 ATP-molekulojn kaj 13 NADPH-molekulojn.
La formado de kolesterolo okazas en pli ol 30 reagoj, kiuj povas esti grupigitaj en pluraj stadioj.
1. Sintezo de mevalona acido.
La unuaj du sintezaj reagoj koincidas kun la ketogeneza reagoj, sed post la sintezo de 3-hidroksi-3-metilglutaryl-ScoA, la enzimo eniras hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA reduktase), formante mevalonan acidon.
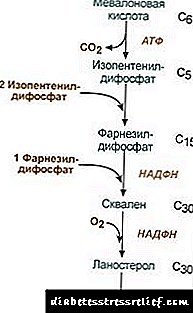 |
Skemo de reago de sinkolesterolo2. Sintezo de izopentenila difosfato. En ĉi tiu stadio, tri fosfataj restaĵoj estas ligitaj al mevalona acido, poste ĝi estas dekarboxilata kaj dehidrogenata. 3. Post kombinado de la tri molekuloj de izopentenila difosfato, farnesil difosfato estas sintezita. 4. La sintezo de skaleno okazas kiam du restaĵoj de farnesil-difosfato estas ligitaj. 5. Post kompleksaj reagoj, lineara skaleno cikliĝas al lanosterolo. 6. Forigo de troaj metilaj grupoj, restarigo kaj izomerigo de la molekulo kondukas al apero de kolesterolo. Reguligo de la agado de hidroksimetilglutaryl-S-CoA-reduktase3. La indico de kolesterola biosintezo ankaŭ dependas de la koncentriĝo specifa portanta proteinoprovizante por la ligado kaj transportado de hidrofobaj interaj sintezaj metabolitoj. Vi povas demandi aŭ lasi vian opinion. La ĉefa punkto de regulado estas la reago de formado de mevalona acido. 1. Alostera regulado. Kolesterolo kaj en la hepato - kaj galaj acidoj inhibicias HMG-CoA-reduktase. 2. Subpremo de sintezo de HMG-CoA reduktase-kolesterolo. 3. Reguligo per fosforilado-defosforilado de HMG-CoA-reduktase, aktiva ne-fosforilata formo. Glucagono kaŭzas senaktivigon, kaj insulino kaŭzas aktivadon tra kompleksa akvofalo de reagoj. Tiel, la indico de sintezoj de kolesterolo ŝanĝiĝas kun ŝanĝo en absorbaj kaj postabsorbaj statoj. 4. La indico de sintezo de HMG-CoA reduktase en la hepato estas submetata al klaraj tagaj fluctuoj: maksimume noktomeze kaj minimume matene. Interŝanĝo de esteroj de kolesterolo En la ĉeloj kolesterola esterigo okazas kiam elmontrita acil-CoA-kolesterolo-aciltransferase (AHAT): Acyl-CoA + Kolesterolo ® NS-CoA + Acylcholesterol En ĉeloj, ĉefe linoleilkolesterolo formiĝas. Esteroj troviĝas ĉefe en la citosol kiel parto de lipidaj gutetoj. La formado de esteroj povas esti konsiderata, unuflanke, kiel mekanismo por forigi troan kolesterolon de la membranoj, kaj aliflanke, kiel mekanismo por stoki kolesterolon en la ĉelo. La mobilizado de rezervoj okazas per partopreno de enzimoj esterasehidrolizantaj kolesterolojn: Acilkolesterolo + H2O ® Grasa Acido + Kolesterolo La sintezo kaj hidrolizo de esteroj estas precipe aktiva en la ĉeloj de la suprena cortekso. En sangaj lipoproteinoj formado de esteroj okazas kun la partopreno de lecitina-kolesterolo-aciltransferasa (LHAT), katalizanta la translokigon de la acila restaĵo de lecitino al kolesterolo. LHAT estas formita en la hepato, sekreciita en la sangofluon kaj ligita al lipoproteinoj. La plej aktiva LHAT en HDL, kie ĝi estas lokalizita en la surfaca tavolo. La esteroj de kolesterolo formitaj ĉi tie estas hidrofobaj kaj trempitaj en la lipidan kernon. En la fosfolipida monostrato estas libera spaco por kolesterolo, kiu povas esti plenigita per kolesterolo el ĉelaj membranoj aŭ el aliaj lipoproteinoj. Tiel, HDL ŝajnas esti kolesterolo kaptita de la agado de LHAT. Bile Acida Sintezo En la hepato, parto de la kolesterolo estas transformita al galaj acidoj. Balaj acidoj povas esti konsiderataj kiel derivaĵoj de kolana acido. Kolanika acido kiel tia ne estas formita en la korpo. En hepatocitoj el kolesterolo, primaraj vezikaj acidoj formiĝas - chenodeoksikolika kaj olika. Post la sekrecio de la galo en la inteston sub la agado de enzimoj de la intesta flaŭro, de ili formiĝas sekundaj galaj acidoj - litokola kaj deoxikola. Ili estas absorbitaj de la intestoj, kun la sango de la portala vejno eniras la hepaton, kaj poste en la galon. Bile enhavas ĉefe konjektajn biliajn acidojn, t.e., iliajn komponaĵojn kun glicino aŭ taurino. La koncentriĝo de biliaj acidoj en galo estas ĉirkaŭ 1%. La ĉefa parto de galaj acidoj okupiĝas pri hepatoenteria cirkulado.Malgranda porcio da biliaj acidoj - ĉirkaŭ 0,5 g ĉiutage - estas elmetita en feĉojn. Ĉi tio kompensas per la sintezo de novaj vezikaj acidoj en la hepato, la fundo bilia acida ĝisdatigas en ĉirkaŭ 10 tagoj. Kolesterolo ankaŭ estas elmetita ĉefe tra la intestoj. Ĝi eniras la intestojn kun manĝo kaj el la hepato kiel parto de galo. Kolesterolo absorbita en la sangon enhavas frakcion devenantan de galo (endogena kolesterolosintezita en la hepato), kaj la frakcio derivita de manĝaĵo (eksogena kolesterolo) Forigo de kolesterolo el histoj okazas per ĝia oksido al galaj acidoj en la hepato, sekvita de ilia ekskrecio per fekloj (proksimume 0,5 g ĉiutage) kaj per ekskrecio de senŝanĝa kolesterolo (ankaŭ kun fekiloj). En stacidomo: (Kolesterolofino + Kolesteroloekz) - (Kolesteroloekskreo + Bileaj acidojekskreo) = 0 Se ĉi tiu ekvilibro ĝeniĝas, la koncentriĝo de kolesterolo en la histoj kaj en la sango ŝanĝiĝas. Pliigita sanga kolesterolo - hipercolesterolemia. Ĉi tio pliigas la verŝajnecon de aterosklerozo kaj galo-malsano. REGULO DE LIPIDOJ Lipida metabolo estas reguligita de la centra nerva sistemo. Longa daŭro negativa emocia streĉo, pliigo de la liberigo de katekolaminoj en la sangofluon povas kaŭzi rimarkindan pezan perdon. Agado glucagono sur la lipolitika sistemo similas la agado de catecolaminoj. Adrenalino kaj noradrenalino pliigi la aktivecon de histo-lipazo kaj la indico de lipolizo en adiposa histo, rezulte pliigas la enhavon de grasaj acidoj en sanga plasmo. Insulino havas la kontraŭan efikon de adrenalino kaj glucagono sur lipolizo kaj mobilizado de grasaj acidoj. Kreska hormono stimulas lipolizon, induktante la sintezon de akenilata ciklase. Pituitaria hipofunkcio kondukas al la deponado de graso en la korpo (pituitaria obezeco). Tiroksino, seksaj hormonojankaŭ influas lipidan metabolon. Forigo de la seksaj glandoj ĉe bestoj kaŭzas troan grason deponado. LIPIDOJ METABOLIKAJ DISORDOJ Kolesterolo estas steroido specifa al bestaj organismoj. La ĉefa loko de ĝia formado en la homa korpo estas la hepato, kie 50% de kolesterolo estas sintezita, 15-20% estas formita en la malgranda intesto, la resto estas sintezita en la haŭto, suprena skalo kaj gonadoj. La fontoj de la formado de la kolesterola fundo kaj la manieroj de ĝia elspezo estas prezentitaj en Fig. 22.1. Fig. 22.1. La formado kaj distribuo de kolesterolo en la korpo. La kolesterolo de la homa korpo (totala kvanto ĉirkaŭ 140 g) povas esti kondiĉe dividita en tri naĝejojn: 30 g), rapide interŝanĝante, konsistas el kolesterolo de la intesta muro, sango-plasmo, hepato kaj aliaj paroksimaj organoj, renoviĝo okazas en 30 tagoj (1 g / tago), 50 g), malrapide interŝanĝante kolesterolon de aliaj organoj kaj histoj, 60 g), tre malrapide interŝanĝita kolesterolo de la medolo spina kaj cerbo, konektiva histo, la ĝisdatiga indico kalkuliĝas tra la jaroj. La sintezo de kolesterolo okazas en la citosol de ĉeloj. Ĉi tiu estas unu el la plej longaj metabolaj vojoj en la homa korpo. Ĝi daŭras en 3 stadioj: la unua finiĝas per formado de mevalona acido, la dua per formado de skaleno (lineara hidrokarbona strukturo konsistanta el 30 karbonaj atomoj). Dum la tria etapo, squaleno estas konvertita al lanosterol-molekulo, tiam estas 20 pluaj reagoj, kiuj konvertas lanosterolon al kolesterolo. En iuj histoj, la hidroksil-grupo de kolesterolo esterigas por formi esterojn. La reago estas katalizita de la intracelula enzimo AHAT (acylCoA: kolesterola acetransferase). La esteriga reago ankaŭ okazas en la sango en HDL, kie troviĝas la enzimo LHAT (lecitino: kolesterola acetransferazo). Kolesteroloj estas la formo, per kiu ĝi estas transportita per sango aŭ deponita en ĉelojn. En la sango ĉirkaŭ 75% de la kolesterolo estas en la formo de esteroj. Kolesterola sintezo estas reguligita influante la agadon kaj kvanton de la ŝlosila enzimo de la procezo - 3-hydroxy-3-metilglutaryl-CoA reductase (HMG-CoA reductase). Ĉi tio atingas du manieroj: 1. Fosforilación / defosforilación de HMG-CoA-reduktase. Insulino stimulas la defosforiladon de HMG-CoA-reduktase, kaj ĝi tradukas ĝin al aktiva stato. Tial en la absorba periodo la sintezo de kolesterolo pliiĝas. Dum ĉi tiu periodo, ankaŭ kreskas la havebleco de la komenca substrato por sintezo, acetil-CoA. Glucagono havas la kontraŭan efikon: per proteino kinase A, ĝi stimulas la fosforiligon de HMG-CoA-reduktasa, igante ĝin neaktiva. Rezulte, la sintezo de kolesterolo en la postabsorba periodo kaj dum fastado estas malhelpita. 2. Malhelpo de sintezo de HMG-CoA-reduktasa.Kolesterolo (fina produkto de la metabola vojo) reduktas la transskriban indicon de la geno HMG-CoA-reduktasa, tiel inhibante sian propran sintezon, kaj galaj acidoj ankaŭ kaŭzas similan efikon. Sango kolesterola transporto estas farita kiel parto de la drogo. LPoj provizas ekzogenan kolesterolon en histoj, determinas ĝiajn fluojn inter organoj kaj ekskrecio de la korpo. Eksogena kolesterolo estas liverita al la hepato kiel parto de postrestinta ChM. Tie kune kun sintezita endogena kolesterolo, ĝi formas komunan fonduson. En hepatocitoj, TAGoj kaj kolesterolo estas enpakitaj en VLDL, kaj en ĉi tiu formo sekretiĝas en la sangon. En sango, VLDL sub la agado de LP-lipase, kiu hidrolizas TAG al glicerol kaj grasaj acidoj, unue estas konvertita al LSPP, kaj poste al LDL enhavanta ĝis 55% de kolesterolo kaj ĝiaj esteroj. LDL estas la ĉefa transporta formo de kolesterolo, per kiu ĝi liveras al histoj (70% de kolesterolo kaj ĝiaj esteroj en la sango estas parto de LDL). LDL el la sango eniras la hepaton (ĝis 75%) kaj aliajn histojn, kiuj havas LDL-receptorojn sur ilia surfaco. Se la kvanto de kolesterolo eniranta la ĉelon superas sian bezonon, tiam la sintezo de LDL-receptoroj estas subpremita, kio malpliigas la fluon de kolesterolo el la sango. Kun malpliigo de la koncentriĝo de senpaga kolesterolo en la ĉelo, kontraŭe, ricevilo-sintezo estas aktivigita. Hormonoj okupiĝas pri regulado de LDL-receptoro-sintezo: insulino, triiodotironino kaj seksaj hormonoj pliigas la formadon de riceviloj, kaj glukokortikoidoj malpliiĝas. En la tiel nomata "transporta reveno de la kolesterolo", i.e. la vojo, kiu certigas revenon de kolesterolo al la hepato, HDL ludas la ĉefan rolon. Ili estas sintezitaj en la hepato en formo de nematuraj pioniroj, kiuj praktike ne enhavas kolesterolon kaj TAG. La pioniroj de HDL en la sango estas saturitaj de kolesterolo, ricevante ĝin de aliaj LP-oj kaj ĉelaj membranoj. La translokigo de kolesterolo al HDL implikas la enzimon LHAT situantan sur ilia surfaco. Ĉi tiu enzimo ligas grasan acidan restaĵon de fosfatidilcolino (lecitino) al kolesterolo. Rezulte, formiĝas hidrofoba molekulo de kolesterolo, kiu moviĝas interne de HDL. Tiel, ne ebriaj HDL, riĉigitaj per kolesterolo, transformiĝu al HDL 3 - maturaj kaj pli grandaj eroj. HDL 3 interŝanĝi esterojn de kolesterolo por TAG enhavitaj en VLDL kaj STD kun la partopreno de specifa proteino, kiu transigas esterojn de kolesterolo inter lipoproteinoj. En ĉi tiu kazo, HDL 3 transformi en HDL2, kies grandeco pliiĝas pro la amasiĝo de TAG. VLDL kaj STDL sub la influo de LP-lipase estas konvertitaj al LDL, kiuj plejparte liveras kolesterolon al la hepato. Malgranda porcio de kolesterolo estas liverata al la hepato de HDL2 kaj HDL. Sintezo de galaj acidoj. En la hepato, 500-700 mg da vezikaj acidoj ĉiutage estas sintezitaj el kolesterolo. Ilia formado inkluzivas reagojn de la enkonduko de hidroksilaj grupoj kun partopreno de hidroksilase kaj reago de parta oksido de la flanka ĉeno de kolesterolo (Fig. 22.2): Fig. 22.2. Skemo de la formado de galaj acidoj. La unua sinteza reago - la formado de 7-a-hydroxycholesterol - estas reguliga. La aktiveco de la enzimo, kiu katalizas ĉi tiun reagon, estas malhelpita de la fina produkto de la pado, biliaj acidoj. Alia reguliga mekanismo estas fosforilado / defosforilado de la enzimo (la fosforilata formo de 7-a-hidroksilase aktivas). Reguligo ankaŭ eblas ŝanĝante la kvanton de la enzimo: kolesterolo induktas transskribon de la geno 7-a-hidroksilase, kaj bilaj acidoj subpremas. Tiroides-hormonoj induktas la sintezon de 7-a-hidroksilase, kaj estrogena repremo. Tia efiko de estrogeno sur la sintezo de galaj acidoj klarigas, kial galia malsano okazas en virinoj 3-4 fojojn pli ofte ol en viroj. Akolaj kaj chenodeoksikolaj acidoj formitaj de kolesterolo estas nomataj "primaraj bilaj acidoj". La plej granda parto de ĉi tiuj acidoj spertas konjugacion - aldonadon de glicino aŭ taurina molekuloj al la karboxila grupo de balea acido. Konjugacio komenciĝas per la formado de la aktiva formo de galaj acidoj - derivaĵoj de CoA, poste taŭro aŭ glicino estas ligitaj, kaj rezulte 4 variantoj de konjugaĵoj estas formitaj: taŭrokolikaj kaj taurohenodeoksikolaj, glukocholikaj kaj glicenodoksikolaj acidaj. Ili estas signife pli fortaj emulsionantoj ol la originalaj bilaj acidoj. Konjugacioj kun glicino estas formitaj 3 fojojn pli ol kun taurino, ĉar la kvanto de taurino en la korpo estas limigita. En la intesto, malgranda kvanto da konjugacioj de primaraj vezikaj acidoj sub la agado de bakteriaj enzimoj estas konvertitaj al sekundaraj vezikaj acidoj. Deoxikola acido, formita de kolika, kaj litokola, formita de deksikolika, estas malpli solvebla kaj pli malrapide sorbas en la intestoj. Ĉirkaŭ 95% de la galaj acidoj, kiuj eniras la inteston, revenas al la hepato tra la portala vejno, poste estas ree sekretigitaj en la galon kaj reuzataj en la emulsifado de grasoj. Ĉi tiu vojo de galaj acidoj estas nomata enterohepata cirkulado. Kun fekcioj, malĉefaj vezikaj acidoj estas plejparte forigitaj. Gallest malsano (kolelitiasis) estas patologia procezo, en kiu ŝtonoj formiĝas en la veziko, kies bazo estas la kolesterolo. La liberigo de kolesterolo en galon devas esti akompanata de proporcia liberigo de galaj acidoj kaj fosfolipidoj, kiuj konservas hidrofobajn kolesterolajn molekulojn en micelara stato. La kaŭzoj kaŭzantaj ŝanĝon en la rilatumo de galaj acidoj kaj kolesterolo en galo estas: nutraĵoj riĉaj en kolesterolo, alt-kaloria nutrado, stagnado de la galo en la veziko, difektita enterohepatika trafiko, difektita sintezo de galaj acidoj, vezika infekto. Ĉe la plej multaj pacientoj kun kolelitiasis, la sintezo de kolesterolo estas pliigita kaj la sintezo de galaj acidoj el ĝi malrapidiĝas, kio kondukas al misproporcio en la nombro de kolesterolo kaj galaj acidoj sekreciitaj en la galon. Rezulte, kolesterolo komencas precipiti en la vezikon, formante viskozan precipitaĵon, kiu iom post iom malmoliĝas. Foje ĝi estas trempita per bilirubino, proteinoj kaj kalciaj saloj. Ŝtonoj povas konsisti nur el kolesterolo (kolesterolaj ŝtonoj) aŭ miksaĵo de kolesterolo, bilirubino, proteinoj kaj kalcio. Kolesterolaj ŝtonoj kutime estas blankaj, kaj miksaj ŝtonoj estas brunaj en malsamaj nuancoj. En la komenca etapo de formado de ŝtono, chenodeoksikola acido povas esti uzata kiel kuracilo. Unufoje en la veziko, ĝi iom post iom dissolvas la kolesterolajn ŝtonojn, sed ĉi tio estas malrapida procezo, kiu daŭras plurajn monatojn. Aterosclerosis estas patologio karakterizita per la apero de aterogenaj plakoj sur la interna surfaco de la vaskula muro. Unu el la ĉefaj kialoj por la disvolviĝo de tia patologio estas malobservo de la ekvilibro inter la konsumado de kolesterolo el manĝaĵo, ĝia sintezo kaj ekskrecio de la korpo. Pacientoj kun aterosklerozo altigis LDL- kaj VLDL-koncentriĝojn. Estas inversa rilato inter HDL-koncentriĝo kaj la probableco disvolvi aterosklerozon. Ĉi tio konformas al la koncepto de funkciado de LDL kiel portantoj de kolesterolo en histoj, kaj HDL de histoj. La baza metala "antaŭkondiĉo" por disvolviĝo de aterosklerozo estas hiperkolesterolemio. (alta kolesterolo en la sango). Hipercolesterolemia disvolviĝas: 1. pro troa konsumado de kolesterolo, karbonhidratoj kaj grasoj, 2. genetika predispozicio konsistanta en heredaj difektoj en la strukturo de receptoroj LDL aŭ apoB-100, same kiel en pliigita sintezo aŭ sekrecio de apoB-100 (kaze de familiara kombinita hiperlipidemio, en kiu koncentriĝas sango kaj kolesterolo kaj TAG). Grava rolo en la mekanismoj de disvolviĝo de aterosklerozo ludas per la modifado de la drogo. Ŝanĝoj en la normala strukturo de lipidoj kaj proteinoj en LDL igas ilin fremdaj al la korpo kaj tial pli alireblaj por kaptiĝo de fagocitoj. Drug-modifo povas okazi per pluraj mekanismoj: 1. glukoksilado de proteinoj, kiu okazas kiam la koncentriĝo de glukozo en la sango pliigas 2. peroksido-modifo, kaŭzanta ŝanĝojn en lipidoj en lipoproteinoj kaj la strukturo de apoB-100, 3. la formado de aŭtoimunaj kompleksoj de la LP-antikorpoj (ŝanĝitaj drogoj povas kaŭzi formadon de aŭtokorpoj). Modifita LDL estas sorbita de makrofagoj. Ĉi tiu procezo ne estas reguligita per la kvanto de kolesterolo absorbita, kiel en la kazo de sia eniro en ĉelojn tra specifaj riceviloj, tial makrofagoj estas superŝarĝitaj de kolesterolo kaj transformiĝas en "ŝaŭmajn ĉelojn", kiuj penetras en la subendotelian spacon. Ĉi tio kondukas al la formado de lipidaj makuloj aŭ strioj en la muro de sangaj glasoj. En ĉi tiu stadio, la vaskula endotelio povas konservi sian strukturon. Kun pliigo en la nombro de ŝaŭmaj ĉeloj, endotelia damaĝo okazas. Damaĝo kontribuas al plateleta aktivado. Rezulte ili sekrecias tromboxanon, kiu stimulas la plaketan agregadon, kaj ankaŭ komencas produkti kresk-faktoron de kresko, kiu stimulas la proliferadon de glataj muskolaj ĉeloj. Ĉi-lastaj migras de la medialo al la interna tavolo de la arteria muro, tiel kontribuante al la kresko de la plako. Plue, la plakaĵo elkreskas kun fibra histo, la ĉeloj sub la fibra membrano estas nekrotikaj, kaj kolesterolo estas deponita en la intercelula spaco. En la lastaj stadioj de disvolviĝo, la plako estas trempita per kalciaj saloj kaj fariĝas tre densa. En la areo de la plako, ofte formiĝas sangoj, blokante la lumon de la vazo, kio kondukas al akra cirkulada perturbo en la responda histo-loko kaj disvolviĝo de koratako. La regulado de la sintezo de kolesterolo - ĝia kerna enzimo (HMG-CoA reductase) efektivigas diversajn manierojn. Fosforilación / defosforilación de HMG-reduktasa. Kun pliigo en la rilatumo insulino / glucagono, ĉi tiu enzimo fosforilas kaj pasas al aktiva stato. La ago de insulino efektiviĝas per 2 enzimoj. HMG-CoA-reduktasa kinasa fosfatasa, kiu igas la kinaseon neaktiva defosforilata stato: Fosfotasa HMG-CoA-reduktasa transformante ĝin en defosforilatan aktivan staton. La rezulto de ĉi tiuj reagoj estas la formado de aktiva formo defosforilata de HMG-CoA-reduktasa. Sekve, dum la absorba periodo, kolesterolo plialtiĝas. Dum ĉi tiu periodo, la havebleco de la komenca substrato por la sintezo de kolesterolo - acetilo - CoA pliiĝas (rezulte de manĝado de manĝaĵoj enhavantaj karbonhidratojn kaj grasojn, ĉar CoA-acetilo formiĝas dum rompo de glukozo kaj grasaj acidoj). En la postabsorba stato, gluĉagono tra proteingenase A stimulas la fosforiligon de HMG - CoA - reduktase, igante ĝin neaktiva stato. Ĉi tiu ago estas plibonigita per la fakto, ke samtempe glukozono stimulas la fosforiladon kaj senaktivigon de la fosfataza HMG-CoA-reduktase, konservante tiel HMG-CoA-reduktase fosforilatan neaktivan staton. Rezulte de tio, la sintezo de kolesterolo en la postabsorbado kaj dum fastado estas malhelpita de endogena sintezo. Se la kolesterola enhavo en manĝaĵo estis alportita al 2%, tiam la sintezo de endogena kolesterolo malpliiĝis akre. Sed la kompleta ĉeso de sintezo de kolesterolo ne okazas. La grado de inhibicio de la biosintezo de kolesterolo sub la influo de kolesterolo venanta de manĝaĵo varias de homo al persono. Ĉi tio indikas la individuecon de la procezoj de formiĝo de kolesterolo. Reduktante la intensecon de kolesterola sintezo, eblas redukti ĝian koncentriĝon en sango. Se la ekvilibro inter la konsumado de kolesterolo kun manĝaĵo kaj ĝia sintezo en la korpo unuflanke kaj la ekskrecio de galaj acidoj kaj kolesterolo aliflanke, la koncentriĝo de kolesterolo en la histoj kaj sango ŝanĝiĝas. La plej gravaj konsekvencoj estas asociitaj kun pliigo de sanga kolesterola koncentriĝo (hipercolesterolemia), dum la verŝajneco disvolvi aterosklerozon kaj kolelitiasis. Familia hipercolesterolemio (HCS) - ĉi tiu formo estas multe pli ofta - ĉirkaŭ 1 paciento po 200 homoj. Heredita difekto en HCS estas malobservo de la absorbo de LDL de ĉeloj, kaj tial malkresko de la indico de LDL-katabolismo. Rezulte la koncentriĝo de LDL en la sango pliiĝas, same kiel kolesterolo, ĉar estas multe da ĝi en LDL. Tial kun HCS estas karakteriza deponejo de kolesterolo en histoj, precipe en haŭto (xanthomoj), en muroj de arterioj. Malhelpo de sintezo de HMG-CoA-reduktase La fina produkto de la metabola vojo de kolesterolo. Ĝi reduktas la ritmon de transskribo de la geno HMG-CoA reduktasa, tiel inhibante sian propran sintezon. La hepato aktive sintezas bilinajn acidojn el kolesterolo, kaj tial la galaj acidoj malhelpas la agadon de la geno HMG-CoA reduktasa. Ĉar HMG-CoA reductase ekzistas post sintezo de ĉirkaŭ 3, inhibicio de la sintezo de ĉi tiu kolesterola enzimo estas efika regulado.
|